
LEARNING OBJECTIVES
After reading this chapter, students should be able to:
- Define what makes a geologic material a fossil
- Summarize the processes that affect organisms in the life to death and preservation transition
- Compare modes of fossilization
- Describe different types of trace fossils
Introduction
Taphonomy is the subdiscipline of paleontology related to the processes of fossilization. This includes all things that happen to the remains of an organism after it dies until it is observed or collected by a geoscientist. This might include the decay of soft tissues, the separation of skeletal hard parts, and chemical changes to the organism after burial. By studying these processes, we can also uncover clues about the environment in which the organism lived, died, and was preserved.
IS THIS A FOSSIL?

Consider the photo of Archaeopteryx on this page. This image likely represents what many people think of when they hear the word “fossil:” bones of an ancient organism preserved in rock. Indeed, it does include major components that define many fossils: easily preservable hard parts (bones), a medium of preservation (surrounding rock), and it’s old. Now think about a turkey leg your dog buries in your backyard. Is that a fossil? No, the turkey leg isn’t a fossil because it is too recent. Officially, fossils are remains of organisms over 11,700 years old, which marks the boundary between the Pleistocene and Holocene Epochs.
_in_Baltic_amber._Age_50_Mill._years_(the_Lower_Eocene).JPG)
Many fossils are body fossils, which includes both mineralized hard parts (shells, bone), and a diverse array of soft tissues (like skin, feathers, flowers) from a previously living organism. Trace fossils (such as burrows, nests, and coprolites) represent the activity of living organisms and are also found throughout the fossil record. Preservation of both body and trace fossils typically occurs in lithified sediment, but body fossils may also be trapped in amber (fossilized tree resin), glacial ice, tar pits, or even lava.
DEATH TO FOSSIL TRANSITION
Decay of soft tissue
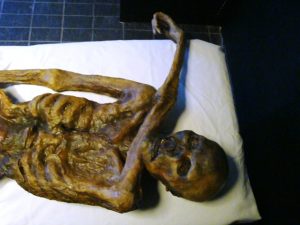
When a living organism dies, soft tissues, as such muscles and internal organs, that make up the body of the organism will typically immediately begin to decay. Scavengers may take advantage of a feast. Bacteria and fungi will begin to break down the tissues to reuse available nutrients. Soft tissue decay happens fastest in the presence of warm, wet conditions with lots of hungry organisms around. Low temperatures, poor oxygen supply, and/or extremely high or low pH slow down rates of soft tissue decay, as these factors inhibit bacterial growth. Therefore, some of the best fossils that include unaltered soft tissues come from deposits that represent rapid burial which sealed the organism off from its environment. Other superb fossils including soft tissue remains are entombed in glacial ice. In some cases a thin film of carbon may be left behind as this rapid burial occurs, leaving a fossil preserved by carbonization.
Pre-burial decay of hard skeletal elements
For multicellular organisms with hard parts (shells, bones, teeth; collectively referred to as skeletal elements), it is the soft tissues that hold these hard pieces together. Once the skin, tendons, muscles, and other connective tissue decays away, there is little to keep the skeletal elements intact, and these shells and bones can disaggregate, becoming sedimentary particles.
_3_(15278777236).jpg)
Most of these skeletal elements are reworked by waves or other currents in the environments in which the organisms lived. If the environment in which the organism lived was prone to currents of running water, flooding, large storms, or turbidity currents, it is possible for the skeletal elements to be transported out of their original habitat.
In high energy environments, such as those with with active currents, it may become very easy for skeletal hard parts to undergo any (if not all) of the following physical processes prior to burial:
- Disarticulation – Separation of valved shells or vertebrate skeletons into individual pieces.
- Fragmentation – Physical breakage of shells or bone into smaller pieces.
- Abrasion – Physical wearing away of shells or bone as they are jostled against each other or against particles of sediment in currents.
Generally, skeletal components that are thin, less dense, and elongate are more susceptible to physical breakage. Chunky, robust, dense skeletal elements are more likely to persist with minimal damage.
There are also some biological processes that can contribute to the physical weakening of skeletal components and increase the surface area for chemical processes to continue breaking down the hard parts, such as:
- Boring – parasitic or predatory organisms create holes via chemical and physical means in shells or bone when the animal is alive. In marine settings, common borers include sponges and snails. After death, additional boring of shells and bones can occur by organisms that require hard substrates for growth.
- Encrustation – organisms that need a hard surface on which to grow may populate the surface of the shells or bone (note for some shells, this may also occur while the shelled organism is still alive). Typical encrusting organisms in marine settings are sponges, bryozoans, tube worms, and barnacles.
Compare the three specimens below–click on the first two to rotate them and consider the differences between the first two well-preserved specimens and the bored and encrusted specimen.
Articulated Mercenaria sp. specimen
![Sponge borings (Entobia made by the genus Cliona) and encrusters on a shell of the modern hard clam, w:Mercenaria mercenaria, from North Carolina. This encrustation and boring happened after the death of the clam when the shell was empty. Photograph taken by Mark A. Wilson (Department of Geology, The College of Wooster), 2007. English Wikipedia [Public domain]](https://opengeology.org/historicalgeology/wp-content/uploads/2019/10/BoredEncrustedShell1.jpg)
Post-burial decay of hard skeletal elements
After skeletal hard parts are buried, additional chemical changes will commonly occur to the shells or bone. Many of these processes are described in the next section on modes of preservation, and are related to the changing the mineralogy of the original hard parts via recrystallization, permineralization, replacement, or dissolution. These chemical changes occur due to groundwater seeping through the pores of the burial sediments, and increasing temperatures associated with deeper burial. As the fossil becomes more deeply buried, additional distortion and fragmentation of the material is possible due to the weight of the overlying sediment. Stretching and shearing of fossils due to applied tectonic stress may also occur.
Death to Fossil Transition
MODES OF FOSSIL PRESERVATION
Unaltered remains
An amazing diversity of fossils exist in a relatively unaltered state. While there may be some compression of the fossil material due to the burden of burial sediments above, or separation of individual shell valves or bones, unaltered remains are skeletal components preserved in their original mineral composition. It is important to have a general sense of what the expected mineralogy of a fossil organism should be to determine whether another mode of preservation could apply to the specimen.
| Common fossil mineralogy | Common organisms |
| Calcite (CaCO3) | Foraminifera, coccolithophores, sponges, tabulate and rugose corals, brachiopods, bryozoans, trilobites, molluscs, echinoderms |
| Aragonite (CaCO3) | Modern (scleractinian) corals, molluscs |
| Quartz (SiO2) | Radiolarians, diatoms, sponges |
| Hydroxyapatite Ca5(PO4)3 | Vertebrate bone and teeth |

In some exceptional cases, soft tissues, such as skin, feathers, or fur may be preserved. This type of preservation does not typically occur within sediments, but rather, in materials like amber or glacial ice.
Permineralization
Permineralization occurs when open spaces within porous tissues are filled in with mineral crystals after the skeletal material of the fossil has been buried. Groundwater containing dissolved minerals passes through the buried fossil, and when that water evaporates, minerals grow in the available space. Typical infilling minerals include quartz, calcite, and iron oxides, which are all common cementing minerals for sedimentary rocks. Permineralization is a common mode of preservation for vertebrate bones and wood, but can also occur in some shells. In the case of petrifaction (“petrified” wood), minerals both fill the porous spaces in the plant tissues and replace the organic material of trees. Permineralized fossils tend to maintain a high degree of detail, as they preserve both internal structures and the three-dimensional shape of the original organism. Because crystal growth occurs in the prior open spaces within the skeletal material, permineralized fossils tend to be denser and heavier than their original counterparts.

Carbonization
When soft tissues are quickly buried in quiet water, they are subjected to increased pressure and rising temperatures as part of the lithification process. These conditions affect the remaining organic material, resulting in a process that releases chemically volatile compounds. When the volatiles are released, a dark-colored film of chemically stable carbon is left behind. Intricate details of delicate soft tissues may be preserved, including individual cell structure in plants, fish scales, insect wings, and even internal organs.
Crane fly, Florissant Fossil Beds National Park
Replacement

Increased pressure and temperature conditions in the deep burial of sediments liberate trapped fluids which contain mobile chemical ions. The fluids interact with buried fossils and may completely dissolve the original mineral or organic material, replacing it with different ions that precipitate from the circulating fluids. Common replacement minerals include silica and pyrite. The decay of organic material requires bacterial support which can assist in reducing sulfur and precipitating the iron sulfide mineral, pyrite. Often, much of the detail of the original organism can be preserved via replacement.
Recrystallization
Recrystallization is an important process that preserves fossils made of aragonite, a common, but chemically unstable mineral over the geologic time. Many marine invertebrates (molluscs, modern corals) have a metabolism that allows them to crystallize aragonite as their skeletal material. Aragonite is a polymorph of CaCO3 and it sometimes appears as beautiful “mother of pearl” on the inside nacreous layer of shells. But outside of the organism’s living tissue, aragonite is less chemically stable than calcite, and so it spontaneous recrystallizes to make calcite after burial. This calcite “version” of the shell is more likely to last long enough to enter the geological record and be discovered by geologists. There is no change to the chemical formula of the mineral during recrystallization, just the same atoms reconfiguring into a more stable crystal structure. This preserves the overall shape of the fossil, but recrystallization can destroy some of the finer details as new crystals form.
Iridescent mother of pearl on nautiloid shell
Internal molds

When the interior space of an organism is buried and filled with sediment that is then lithified, the interior shape is preserved as an internal mold. Imagine making a sand castle at the beach, filling a plastic bucket with wet sand, then inverting it onto the ground and lifting the bucket away. The shape of the inside of the bucket is preserved though the bucket itself is gone–that’s an internal mold. In geology, this mode of preservation is usually found in bivalves or brachiopods, or snails that have a single open chamber where sediment can get in and work its way into the entire space. This mode of preservation can reveal lovely impressions of internal features like muscle scars, but since most molluscs are more readily identified using external ornamentation, loss of the original shell can make things tricky for paleontologists.
In some cases, small cavities inside a shell may remain after sediment is washed into the shell. The sediment sinks to the bottom of the cavity and leaves a pocket of water above. Over time, these pockets can later close up with precipitated minerals, potentially serving as geopetal structures. Geopetal structures are “up indicators”, and give geologists a good sense of which way was up when the bed was deposited, which can be helpful telling the stories of sedimentary layers that have been deformed by tectonic processes. In the image below, examine the difference in sizes of the infilled calcite crystals in the area outlined in pink versus the area outlined in orange. Which way was up?
Geopetal structures in Reynolds Limestone
External molds

An external mold is an impression of an organism’s outside shape. This type of preservation commonly occurs when a fossil is pressed into soft sediment. The original hard parts of the organism may then be transported away by currents, or after burial and lithification, the original hard parts may dissolve away. When external molds are discovered, the observer sees a version of the exterior of that organism, but it is not an exact replica because it shows negative relief. Negative relief means that on the impression, raised areas represent indentations on the original fossil, while depressed areas represent raised areas. If you have ever walked along a beach, turned, and examined the footprints you left behind, these are negative impressions of your feet. If that sand could lithify instantly, it would represent an external mold of your foot.

Fossil trackways, a type of trace fossil, are a type of external mold. Some very famous dinosaur tracks are preserved in this way. Most often, external molds are created by hard shells or bone. However, some external molds of dinosaur skin have been discovered, meaning that even soft tissues can be preserved via this mode under the right conditions.
Another method of creating external molds is through bioimmuration. Bioimmuration comes in several forms, but consists of the creation of an external mold through the encrustation of other organisms on exterior surfaces.

For example, barnacles often encrust boats and if you were to remove the wood from the barnacle, an impression of the wood would remain on the bottom of the barnacle. Sometimes, whole successions of external encrusters will attach themselves. These can certainly include arthropods such as barnacles, but also encrusting corals, brachiopods, sponges, and many more organisms just looking for a place to attach. Bioimmuration as a form of external mold only happens when the original hard substrate dissolves, rots, or is otherwise removed, leaving behind the encrusting organisms and, very often, a history of successive encrustations that give important glimpses into unique paleoecological situations.
External mold (left) of a trilobite body fossil in a concretion
Casts

Casts are replicas of organisms, essentially recreating their original shape. First, an organism must be preserved as an external mold, or perhaps open burrows preserved as trace fossils. The impressions or other cavities are then filled in with sediment, producing a three-dimensional form that mimics the form of the original fossil. Imagine if you took some clay and pressed it into the external mold of the trilobite above. Once hardened, you could pop that trilobite out and have a cast of the original fossil, mimicking its original form, and presented in positive relief. Some details are typically lost when a cast is created, but can still provide critical information about the organism. Interestingly, many fossil skeletons on display at museums are primarily casts. This doesn’t mean the fossils are fakes since the casts are created based off the original fossils. Casts are used for many reasons: the original fossils can be hundreds of pounds of permineralized bone making it a challenge to mount the specimens, the originals may be fragile and require a controlled environment for preservation, it is a way to share specimens among museums, or the fossils may be currently be under study.
Modes of Fossil Preservation
TRACE FOSSILS: STUDIES IN SCURRYING, SCRAPING, AND SLITHERING
As noted earlier, remains of organisms are not just found as body fossils. Trace fossils can yield interesting perspectives on how an organism lived and interacted with its environment. After all, if someone excavated your fossilized bones, would that tell your entire story? Would your bones and how were buried say anything about what you were doing at any point in time? Likewise, non-human organisms scurry, burrow, step, jump, run, walk, slither, and grow in a variety of ways. Traces of these actions are preserved in sediment and are studied under the branch of paleontology called ichnology.
Earliest trace fossils
Some of the earliest trace fossils are found in rocks that are around 3.5 billion years old, and preserved as structures referred to as stromatolites. These are macroscopic structures that have a dome-like shape. In them, wafer-thin mats of photosynthetic microbial mats (cyanobacteria) alternate with fine calcite mud. Initially flat, they grow increasingly convex through time, though the relief and size vary tremendously. Stromatolites require certain conditions to grow: (1) sunlight, and therefore shallow water, and (2) absence of grazing predators such as snails. They are still forming today in places like Shark Bay of western Australia and tidal channels in the Bahamas, both locations where predatory grazing by snails is absent. The high salinities and powerful scouring tides of these environments generally keep the snails away. The cyanobacteria that made the ancient stromatolites are not preserved, but are evidenced by the layers of sediment that stuck to the cyanobacterial mats. Since the Archean, evolution has led to more complex organisms using ever-changing forms of locomotion. New and unique forms of trace fossils emerged over time including horizontal and vertical burrows, and eventually, footprints.
Classification of trace fossils
Trace fossils are often classified by an ecological method that focuses on the behavior of the organism (see table).
| Trace fossil classifications (after Seilacher, 1967) | ||
| Ethologic (behavior) class | Associated behavior | Example |
| Repichnia | Locomotion |
 |
| Cubichnia | Resting |
 |
| Domichnia | Dwelling |
 |
| Pascichnia | Grazing |
 |
| Fodinichnia | Feeding burrows |
 |
Note: Coprolites, or fossilized feces, are also considered traces but do not have a formal classification.

An alternative trace fossil classification is similar to the Linnaean system, creating an ichnotaxa from an ichnospecies within an ichnogenus. An example of such an ichnotaxa is Skolithos linearis–a long, straight, vertical burrow. Over time, ichnologists, or paleontologists who study trace fossils, have been able to show relationships among traces and how those traces change shape over time, using cladistic methods similar to those applied to living organisms.Unfortunately, it is usually very difficult to associate a particular organism with a particular t race. However, in rare instances, organisms such as trilobites have been preserved within a trace they created (e.g., Cruziana sp.). The real utility of trace fossils is in their association with particular environments.
Some traces are found in beach facies. Some traces are characteristic of beaches, while other different traces are typical of deep-water muds, and yet others are found in terrestrial mudflats. Even though the shape and movements of particular organisms play a role in how a trace fossil is formed, the characteristics of the sediment are just as critical to creating traces. Using uniformitarianism, geoscientists can use the tools of “neoichnology”, or the study of modern traces, to help understand traces found in rocks and to interpret their likely environment of formation. Importantly, many traces are time-independent. Organisms burrow now just as they did 400 million years ago; though the organisms may be different, the action is the same (Seilacher, 1967). Thus, trace fossils are excellent facies indicators and provide fabulous insights into the organisms and environments that formed them, and have been grouped into categories known as ichnofacies. Each ichnofacies is created by a community of ichnotaxa.
Trace Fossils
Further reading:
University of California Museum of Paleontology Fossils
Seilacher, A. (1967). “Bathymetry of trace fossils”. Marine Geology. 5: 413–428. doi:10.1016/0025-3227(67)90051-5.