John Garrett image, CC BY-SA 3.0, Public domain image via Wikimedia.
LEARNING OBJECTIVES
At the end of this chapter, students should be able to:
- Define biological evolution.
- Distinguish between macro and microevolution.
- Distinguish between different patterns that occur in nature as a result of evolutionary processes.
- Describe how life is organized using systematics.
- Identify some of the key ideas and players in the history of evolutionary theory.
- Identify the key paradigms in the development of evolutionary theory.
- Identify the ways that genes are passed on through various forms of inheritance to the next generation.
Evolution: Patterns in the Biosphere
The biosphere is replete with patterns. This is true not only in the ecosystems around us today, but also in our planet’s fossil past. Some of these patterns include some of the basic things we learn about in introductory biology courses. These patterns force scientists to ask questions. Why are ecosystems arranged in a hierarchy with primary producers on the bottom and omnivorous organisms on top? Why is there so much shared DNA among life forms that seem so different, even across different environments? Why do so many life forms that live such different lives have such similar features? Why do humans and reptiles share four limbs? Why do both butterflies and birds have wings? Why do some features of organisms not have clear functions? Why do I need to put on a jacket, hat, and gloves to venture out into the winter cold?
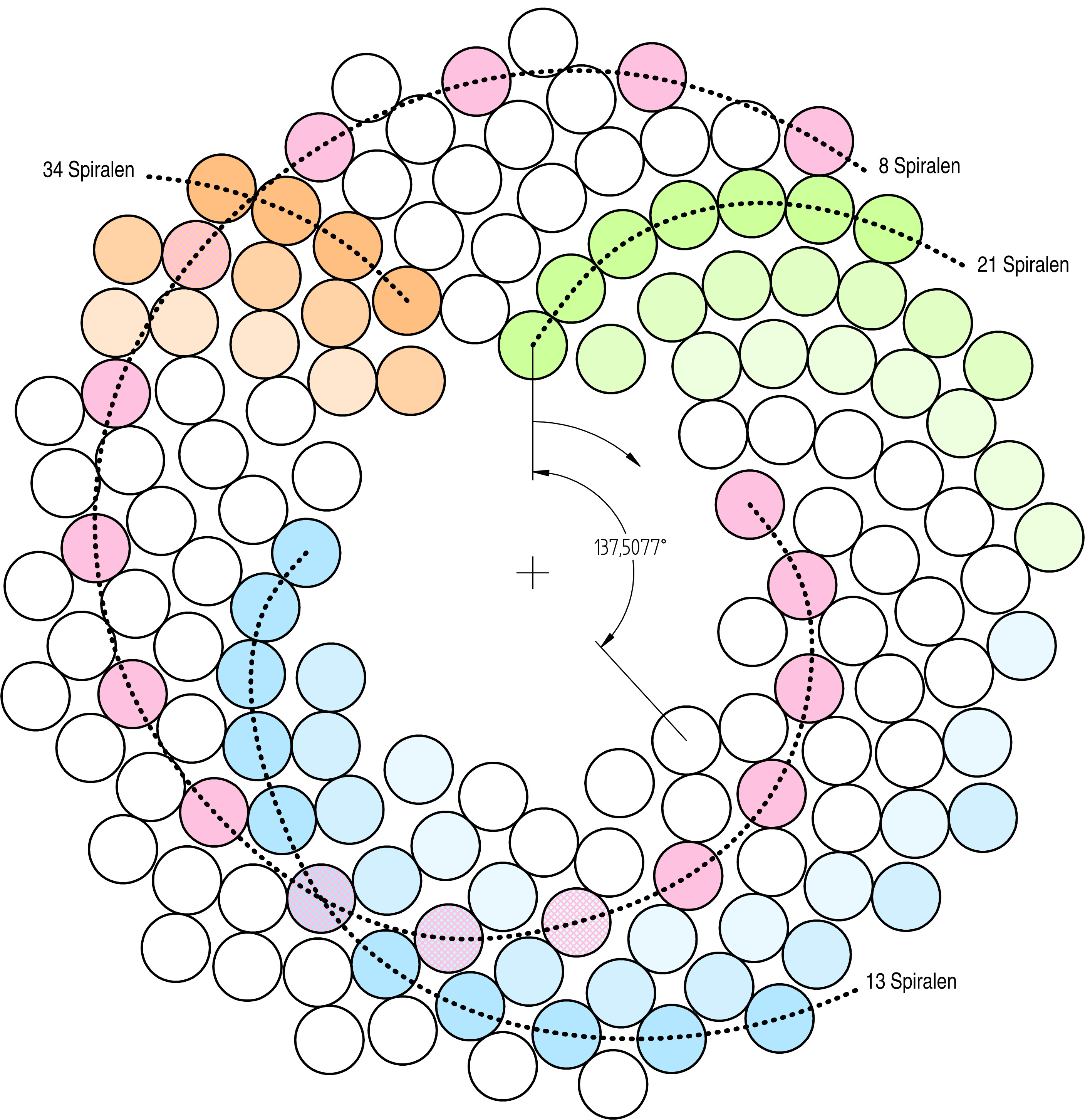
One common pattern in the living world is governed by the mathematical Fibonacci Sequence. This simple sequence of numbers describes an amazing variety of phenomena, with names like the “golden ratio” and the “golden spiral.” It can also be used to describe breeding patterns in organisms and subsequent geometric population growth. A Fibonacci Sequence describes a doubling with each calculation. Over time, it is possible to use this simple relationship to predict the number of organisms present after any number of generations, if resources are available and population growth is unchecked. Also predicted by this equation are other natural patterns, like the Fibonacci Spiral seen in the images below., The arrangement of seeds in a sunflower and the whorls of an ammonite are excellent examples of how this mathematical relationship governs some biological growth patterns. These spiral patterns are based upon a curve that, while growing, keeps its shape and central angle at all scales.

By John Garrett , CC BY-SA 3.0, (Source: Wikimedia Commons, Anna Benczur, CCA ASA 4.0).
By John Garrett – https://www.skepticalscience.com/print.php?n=1959, CC BY-SA 3.0, https://commons.wikimedia.org/w/index.php?curid=74327875(Wikimedia Commons, Andree Stephan, CC ACA 3.0)
Gigapan of fossils accumulated on the Cretaceous Interior Seaway seafloor at Fort Hays State University (Image: Ron Schott). Can you see fossils that display distinct patterns? Do you see a golden spiral emerge?
Evolutionary theory presents us with answers to these questions. Using it as a framework, we can answer these questions and more. Evolution explains the observable patterns in the living world around us. It also explains the patterns we see in the fossil past.
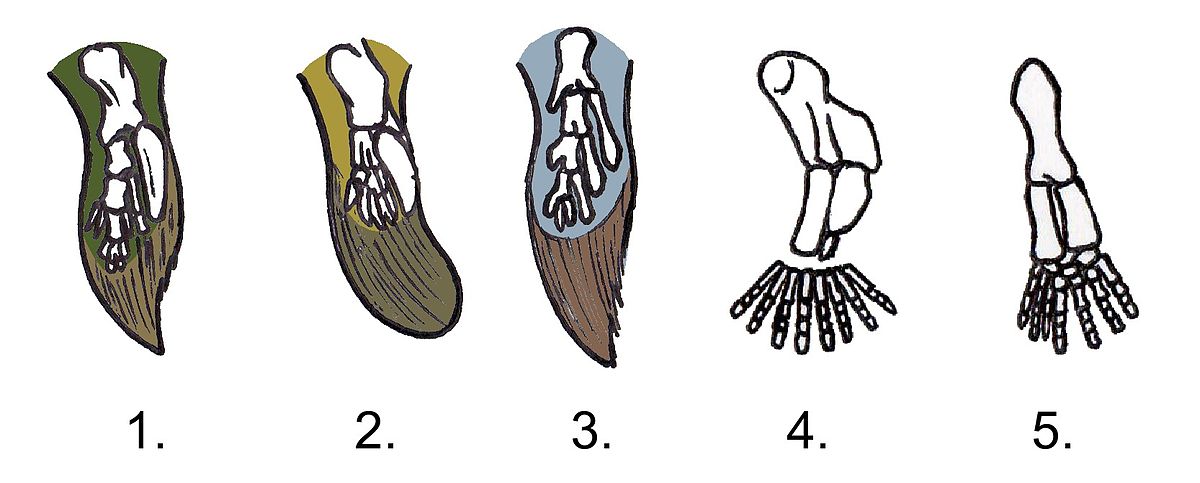
Evolution means “change over time.” It is used in many ways and in many fields. Biological evolution is the mutation of genetic material through generations of offspring. Such evolution from one generation to another does not follow a trajectory. It is the result of the accumulation of changes in DNA. Such modifications occur at different rates in different groups of organisms and at different moments in time, but they are only observable when mutations in an organism’s DNA, its genotype, are physically expressed through its phenotype, or appearance. Such changes in genes can lead to longer horns, more toes, improved leg bone structure for walking on dry land, and bipedalism in primates. These are just a few examples. Billions of years ago, when microbes were the only living things on the planet, such exotic future modifications were not inevitable or predictable. Our own species, Homo sapiens was never guaranteed existence. Still, we are here, and we are able to contemplate questions of origins. You are reading this chapter, and that is a result of evolutionary change! Ultimately, evolution tells us that we are all related. Yes, humans are related to one another, but humans are also related to sponges, flowering plants, and bacteria. All life on this planet is descended from a common ancestor, and was then teased out into millions of distinct evolutionary trajectories. Some of these probably look and act pretty similar to the most ancient ancestral forms of life (their features are said to be evolutionarily “conserved”), and some are extremely different – they are “highly derived.” Descent with modification is the essential idea of evolution.

By John Garrett – (Wikimedia Commons, Thompsma CC BY 3.0) .
Biological evolution helps us understand the living world around us. It helps us understand ourselves as a species. It helps us explain the patterns we see in fossils contained in rocks. Evolutionary change is observable everywhere. The development of antibiotic resistance among bacteria. A stroll through a wooded area will reveal an ecosystem of food webs where an intricate daily dance is performed between organisms that have evolved together over time. A decomposing organism is broken down by other organisms specially adapted for that purpose. Parasites exist within your body, your skin, that you rely on for life but yet never meet as you would a friend or neighbor. Evolution can explain all of this and more. Because it is still occurring today, we can observe its processes in action. As a theory that has been itself evolving for more than a century, it is built upon a robust foundation of evidence from the fossil record, biochemistry, observations of naturalists, and observations still being made in labs, remote tropical forests, and even on other planets. Evolutionary theory binds all of these threads of evidence together into a coherent view of life.
“There is grandeur in this view of life, with its several powers, having been originally breathed into a few forms or into one; and that, whilst this planet has gone cycling on according to the fixed law of gravity, from so simple a beginning endless forms most beautiful and most wonderful have been, and are being, evolved.”
– Charles Darwin, The Origin of Species (1859)
Systematics: Organizing Life
Macroevolution and Microevolution
The fossil record provides excellent evidence for the history of life. In particular, fossils reveal macroevolution, or how species have evolved over time. Microevolutionary change, or changes in gene frequency on very small scales, also occurred in fossils when they were alive. However, fossils do not preserve such change. Macroevolutionary change recorded by fossils shows visible variation in both physical features (body fossils) and behaviors (trace fossils), as well as the introduction of new organisms. We find that not only is all life related, but that life has evolved, through common biochemical means, over the course of more than four billion years. This evidence allows us to build models for how evolution occurred in the past and insights into how it is still occurring today. One such model is the “Tree of Life.” Speciation, or origination, happens at different scales of time, depending on the time it takes to produce offspring. It happens very quickly in bacteria, but quite slowly in larger animals. Because macroevolutionary change can take a great deal of time, the best place to look for evidence of this change in the fossil record. Macroevolution can be thought of as the accumulation of microevolutionary changes, which include various kinds of genetic mutation processes and trait selection. These processes operate at much smaller scales and do not get recorded in fossilization. Despite not being visible in the fossil record, we can study how they work today, and apply uniformitarian principles to understand how they must have worked in the past.
The OneZoom “Tree of Life Explorer”
Linnaeus and His Method
People have long organized and classified organisms. The beginnings of recorded classification go back to Aristotle, with his observations of invertebrates while living on the islands of Greece. His method of classifying organisms began by separating animals from plants. He would go on to separate animals into two further groups that we still use loosely today, Anhaima (animals without blood, or invertebrates) and Enhaima (animals with blood, or vertebrates). This system would persist, like so much of Aristotelian thought, through the middle ages and right up to the time of Linnaeus. Classification of organisms in the years just prior to Linnaeaus’ work took place at a frenzied pace, with new specimens constantly showing up in Europe from the far corners of the world. The “Age of Exploration” (known in the non-western world as the “Age of Being Explored by Europeans”) was a time of great discovery, but each new discovery was as likely to bring confusion as confirmation. Classification was very disorganized, with resulting names of organisms becoming very long and inconsistent. By introducing the system of binomial nomenclature, which we still use today (Genus followed by species, e.g. Homo sapiens), Carl von Linné simplified and unified taxonomy. The system relies on shared characteristics, by analyzing what organisms have in common in order to create descriptive names and then organizing them into groups or “bins.”
Ultimately, understanding the complex cacophony of life is challenging. If evolutionary changes occur in predictable ways, then it should be possible to organize all of this into a “model” of life. Modern science owes its understanding of evolutionary processes to the earlier work of luminaries like Linnaeus, who tried to provide order to the living world around him. His approach to taxonomy, the scientific classification of organisms, is a nested hierarchy, with “Kingdoms” as at the highest taxonomic rank and the most inclusive category. Kingdoms get divided up into “Phyla,” then “Classes,” “Orders,” “Families,” “Genus,” and finally “Species.” Like many students, you may have once created a mnemonic device when you first learned this in school. In 1990, Carl Woese would add a higher taxonomic rank, the Domain, because of the discovery of three distinct forms of RNA that separate the archaebacteria, Eukarya, and Bacteria from one another (Figure below). Other than this change, The system has remained largely unchanged since Linnaeus created it. Though Linnaeus only used anatomy as the basis of his classification decisions, his scheme has largely been validated through insights later derived from DNA sequencing. In a few cases, small refinements have been provided through genetic insights, but overall, Linnaeus’ system provides clarity for the patterns we see!
(Wikipedia Commons)
Lamarckism and Evolutionary Foundations
The theory of evolution provides a robust model for how life changes over time that allows scientists to not only describe what they observe occurring, but to also test ideas about processes related to speciation, genetic mutation, and similar important topics. However, descent with modification is just a part of the story. Determining a mechanism for understanding why some “modifications” helped create success and others did not would rely on uniting these ideas with the environment in which an organism lived. In late 18th century France, Jean-Baptiste Lamarck tried to explain such observations by attributing parents with the ability to pass on traits or behaviors through use or disuse of an organ, anatomical feature, or behavior picked up during their lifetime. This late 18th century idea would become quite a popular way to explain the length of a giraffe’s neck, for instance, which would become progressively longer during its lifetime as it strained to reach high branches. This long neck in the parent could then be inherited by the giraffe’s offspring. Continued use of this trait would enable further development of the feature. Overall, Lamarck viewed evolution as trending from more simple to more complex forms, because he believed in a naturalist origin of life through spontaneous generation. New life forms, such as microbes, evolved toward more complexity through the generations as they adapted to their environment. His ideas were a major step forward for two reasons. He correctly argued that life took its form through adaptation to the environment, an idea that was quite controversial at the time. Though aspects of his ideas would later be proven wrong, the concept of traits being inherited by subsequent generations through natural processes would be a major step forward.

(Wikipedia Commons)
Revolutionary Ideas: Darwin and Wallace

(Wikipedia Commons)
A New Paradigm Emerges: Darwin and Wallace
Scientific discovery often occurs over time, but always occurs through the accumulation of data. Thomas Kuhn (1962) outlined how scientific “revolutions” are structured as changes in a paradigm. A paradigm is a pattern of thinking. Old models and theories give way and evolve as new information arises. Sometimes, the entire pattern of thinking shifts in a major way, marking an entirely new way of approaching scientific problems. Charles Darwin and Alfred Russel Wallace both discovered, pretty much at the same time, the key process that would lead to a new paradigm in evolution. This key process was natural selection, the variation in survival due to advantages and disadvantages in certain traits within an organism. Before Darwin and Wallace, Linnaeus’ ideas showed the relatedness of life and Lamarck’s ideas about inheritance supposed that life has changed over time. These ideas represented the paradigm of the time. Darwin’s publication of “The Origin of Species” in 1859 initiated a major shift in the view of how traits are transmitted from parent to offspring. Rather than parents passing traits directly to offspring through use or disuse, Darwin and Wallace would argue that this occurred through natural selection. Their views on this were influenced by two major factors. First, Thomas Malthus’s writings on economic ideas regarding population and resources emphasized that population growth eventually far outstrips available resources in an environment. Second, both Darwin and Wallace had the opportunity to conduct extensive fieldwork in locations filled with biodiversity relatively untouched by intensive human development.
Charles Darwin famously spent five years (1831-1836) as a naturalist aboard H.M.S. Beagle, a British ship with a mission to circumnavigate the globe. During this time, Darwin was able to spend extended periods of time on land, exploring regions around the South American coast, from Brazil to the Galapagos and beyond. He collected rocks, fossils, animal specimens, planet specimens, and more. Sending them on to England prior to his return, these collections made him famous and would become his life’s work. Ultimately, the immense variation in plant and animal species across South America was puzzling. He found many completely different species living in environments that were very similar. Why was there such variation across distances if the environments were so similar? The classic example of this are his observations of finch variation across the Galapagos Islands (Darwin’s Finches). The best way to characterize this variety was by invoking a common ancestor sometime in the past that, as its descendants moved from island to island, evolved into new species. Another example from his voyage came from his explorations of the greater rhea and lesser rhea, two flightless birds that local guides often ate as food in the area around Bahia Blanca, Patagonia. These two birds were very similar, but different species. Because one lived north of the Rio Negro (River) and the other south, their ancestors must have pursued separate evolutionary trajectories. Darwin’s insights were not relegated only to living specimens. Darwin also collected many fossils. In subsequent study back home, he made connections between his living specimens and fossil ancestors. Such evidence would become very important for the development of his ideas around natural selection.

(Wikipedia Commons)
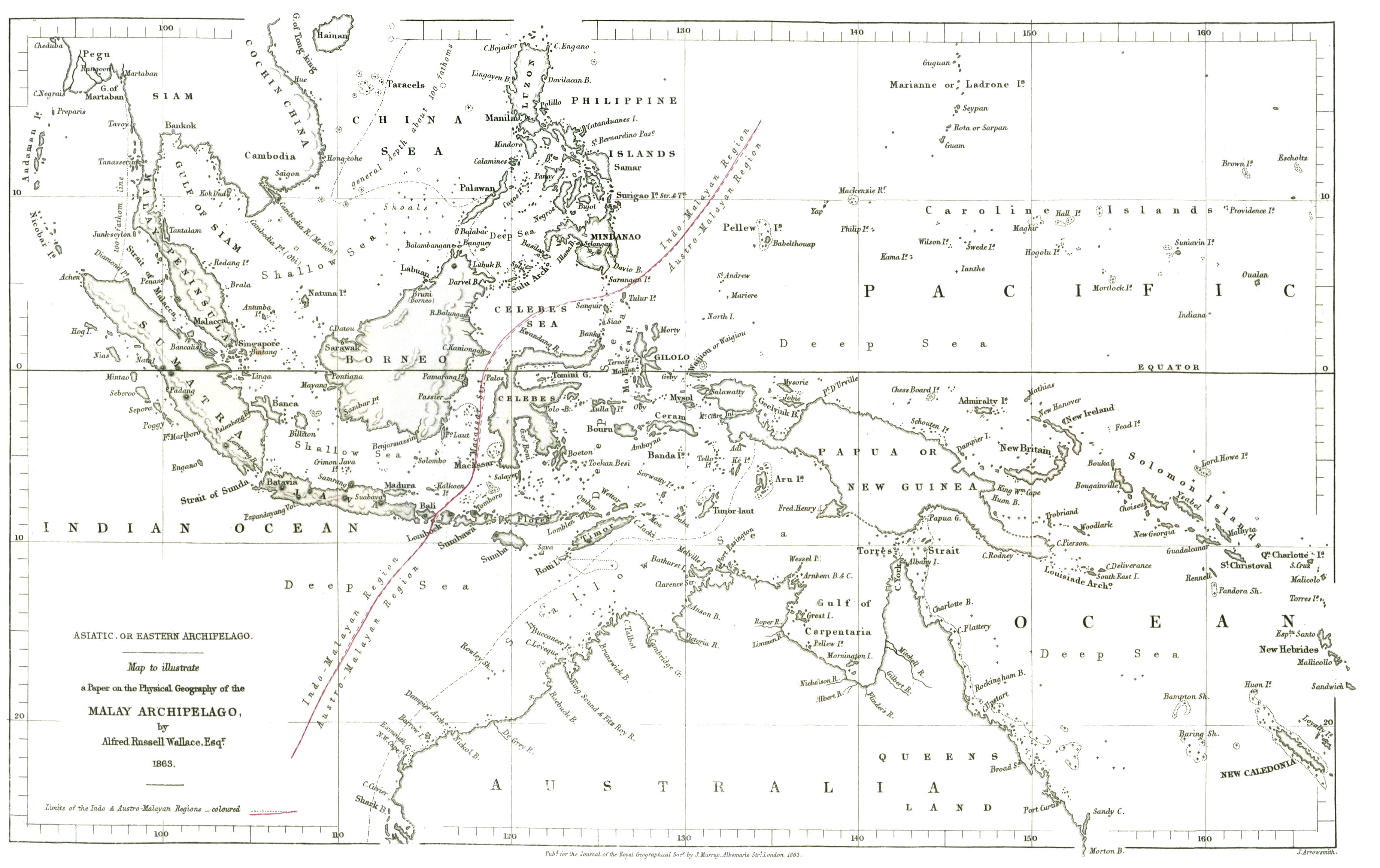
Alfred Russel Wallace conducted most of his work in two places. Like Darwin, he spent a great deal of time in South America, particularly Brazil. He also conducted field work in what is today Malaysia, Indonesia, and nearby places. Inspired by the field work of Darwin and others, he funded his expeditions, running between 1848 and 1862, through the sales of collected specimens. He methodically planned his field work to explore ideas he had surrounding biogeography, or the geographic distribution of species. If evolution occurs, he reasoned, closely related species should live close to one another. His field work played this out, noting that rivers and other bodies of water did indeed separate related but distinct species. His classic example, called the “Wallace Line,” notes the separation of species of Asian and Australasian descent by the Sunda Straits.
(Wikipedia Commons)
He noted that while some species likely descended from a common ancestor had evolved very differently on either side, certain species of flora were the same on both sides. These straits being about 35 km in width, the mobility of an organism was key. Plant pollen and seeds, blown by the wind or transported by water, allow growth on both sides of the strait. Large mammals and most birds, by contrast, could not traverse it. This separation, over time, would lead to new species where once an ancestral form existed. At the time, Wallace would not have known what we now know about the role of plate tectonics in this biogeographical separation. But, he was able to correctly observe here, and elsewhere in his work, that biological change occurs as new species arise from prior species. He recognized natural selection as the mechanism for these observations. Favorable traits, or traits that gave organisms an adaptive advantage, were retained while unfavorable traits would make organisms unfit for their environments.
The two men did correspond. While Darwin came to the idea before Wallace, it was in conversation with Wallace that natural selection, and the new paradigm that would follow, emerged. In 1858, they jointly read papers at a meeting of the Linnaean Society, which would mark the first public description of evolution through natural selection. This was followed one year later by Darwin’s publication, On the Origin of Species. Darwin and Wallace described natural selection as the mechanism that makes survival to the next generation possible, though it does not mean that these offspring are necessarily the “fittest” to survive. Organisms just have to have enough favorable characteristics to be able to survive long enough to pass on their genes, in whatever condition, on to the next generation. Sometimes, genetic mutations passed on are benign. Other genetic changes passed on provide a favorable adaptation for offspring. At still other times, mutations passed on are detrimental to the survival of offspring or even the ability of a parent to reproduce at all.
Meanwhile, Back in Europe: Mendelian Inheritance and Genetics Take Shape
Even with this new understanding of natural selection, it would take an understanding of how genes work to explain how they are inherited by offspring. The foundations for understanding inheritance would come from the 1860s work of an isolated monk in a Czech monastery. While Darwin and Wallace toiled around the globe and in the heat of rainforests, an important part of evolutionary theory was taking shape through humble pea plants. Gregor Mendel, the son of a farmer, was very interested in plants. Harnessing his curiosity and focusing on pea plants, he crossbred a wide array of varieties and recorded how traits were passed down in the next generation. Applying his mathematics training, Mendel was able to use statistics, applied to his pea plant populations, to predict which traits – smooth skin, wrinkled skin, and so on, would be inherited. As is often the case with new scientific insights, this work was not recognized for its importance right away. But, owing to the work, interests, and curiosity of this isolated monk, natural selection was bolstered with genetics. Natural selection would not only be attributable to environmental factors, but could now also be described by changes in how genes are inherited from one generation to the next.

(Wikipedia Commons)

(Wikipedia Commons)
There are three principles in Mendelian genetics. It is also worthy to note that, though the term “Laws” here are accepted within the scientific community, there is enough deviation from them that they may be more accurately referred to as “Principles.” Genes come in two forms called alleles (“Law of Segregation”). Alleles can be dominant (expressed in an organism’s appearance) and recessive (not expressed). This is the “Law of Dominance.” You may remember creating Punnett Squares in a previous biology class –these are tools that can be used to analyze the expression of one allele over another from one generation to the next. The selection of one allele of a gene over another, however, occurs independently of selection occurring among other genes (“Law of Independent Assortment”). These laws, however, do not describe how one allele is transmitted to the next generation, only that alleles exist, are treated independently of other genes during selection, and ultimately determine what variants of such genes are physically expressed. For instance, there are cases where one allele is not completely dominant over another, a situation called “incomplete dominance.” Likewise, there are situations where the phenotype for both alleles is expressed. This is called “codominance.” It is also known that some genes exist in nature with more than two alleles, or “multiple alleles.” Finally, some traits are actually “cogenetic,” such as the color of fruit fly eyes, where several genes contribute to a physically expressed variation.
Mutation, Migration, Drift, and Natural Selection
There are several key processes that drive evolutionary change at all scales. These are genetic mutation, gene migration, genetic drift, horizontal gene transfer, and natural selection.
Genetic mutations are simply changes in the sequence of nucleic bases in the coding portion of a DNA molecule. These can be caused by errors in replication or repair of a sequence as it is passed on to the next generation.
Gene migration, or gene flow works differently. Sometimes, a new, sexually reproducing, individual from a separate population is introduced into a new population of the same species. Introduced traits can include many things, such as new varieties of colors within an insect population. This new and unique genetic material mixes into the descendant populations and can then lead to unique and novel down-generation changes. Genetic material can also be exchanged with the donor population in the same manner as populations subject to gene migration are not necessarily isolated.
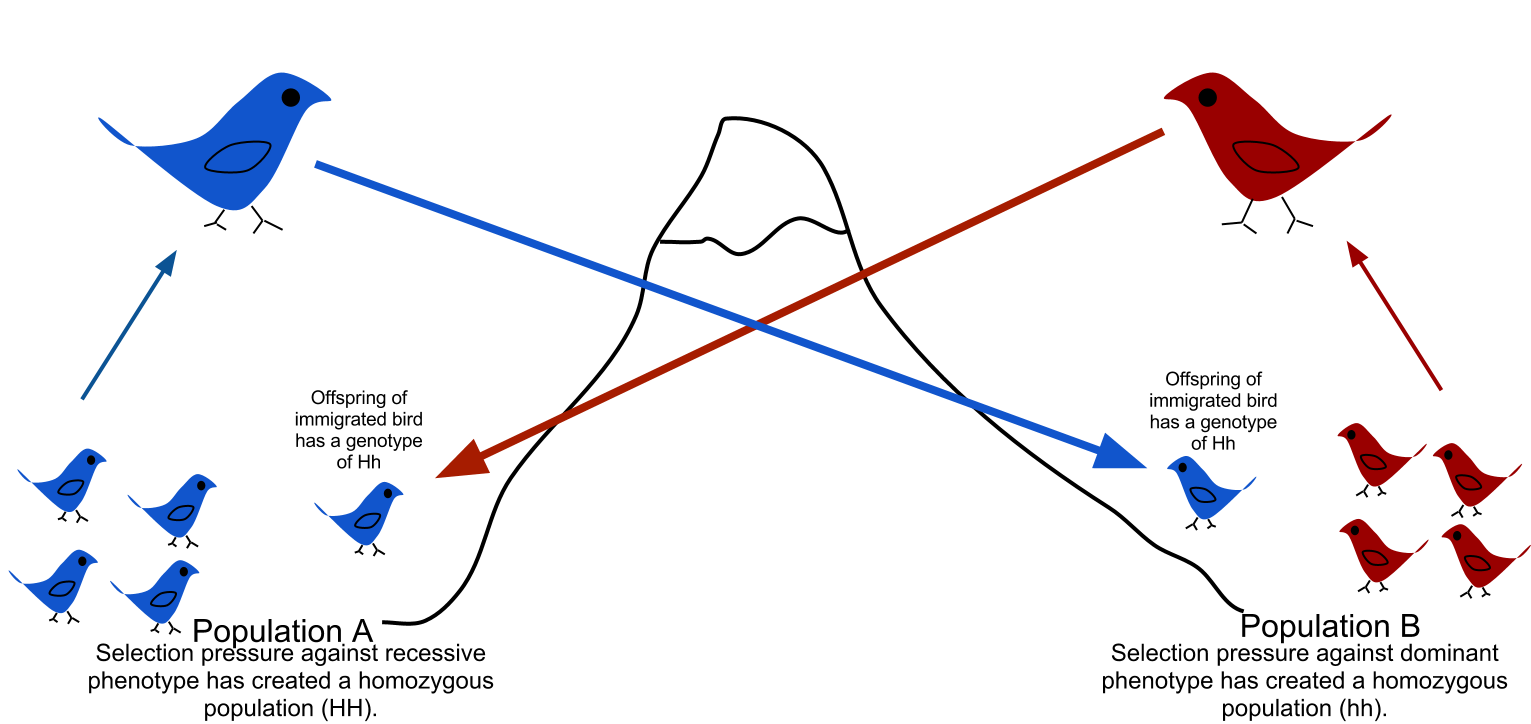
(Jessica Krueger, Wikimedia Commons CC BY-SA 3.0)
Genetic Drift does not lead to adaptational changes in a population. Rather, it is the result of chance, most commonly because some individuals have left behind more offspring than others. It is a change in the frequency of an allele over time, and is completely independent of environmental changes. The genes of the following generations were passed on by those of the individuals fortunate enough to reproduce successfully, and not necessarily of the “fittest.” One simple example comes in populations reduced to small numbers that then recover, such as the American Bison, which was nearly hunted to extinction. While its population is much larger than in the past, its genetic variation is much lower than it was 200 years ago (Ungerer et al., 2012). A simpler example might come among a human couple, each with different eye colors, say brown and blue, but where brown represents the dominant allele. Even if the chance of having brown eyes is 50%, all children may statistically end up with blue eyes, thereby eventually erasing that dominant allele. Such cases provide excellent examples of where phrases like “survival of the fittest” are at best an oversimplification of evolutionary processes.

(Wikipedia Commons)
Two other important evolutionary mechanisms that also occur at the genetic level, but in different ways, are horizontal gene migration and genetic symbiosis. Gene transfer, as we usually think of it, is vertical. This means that genetic material is transferred from parent to offspring. Horizontal gene transfer occurs sideways (Figure left), as with viral transfer. It also occurs among bacteria and archaea that have no means of sexual reproduction. Most of us are familiar with viruses, as we suffer from them frequently and, if we are responsible and had responsible parents, have been vaccinated against them wherever possible. Bacteriophages are a kind of virus that replicates within bacteria and holds promise toward fighting antibiotic resistance (Bragg et al., 2014). Because they inject their genome into host cells where it can replicate, viruses are able to transfer DNA from related and unrelated organisms. This method of transfer is called transduction. Plasmids, which most often are transferred horizontally via a process called transformation, are genetic material that exists in a cell independent of chromosomal interaction. Transformation is the transfer of genetic material between cells. The final method of horizontal transfer, which can occur via the transfer of plasmids or transposons (a chromosomal segment that can undergo a change of location), is conjugation, which occurs through direct physical contact between cells. Genes can also be transferred horizontally through gene symbiosis, which results from close ecosystem interactions between, at times, species that are evolutionarily separated a great deal. Endosymbiosis is thought to be the origin of key eukaryotic organelles such as mitochondria and chloroplasts (each of which brought its own DNA with it into the host cell). Fungi may transfer genetic material to an arthropod, such as an aphid, through exchanges of cellular material from close physical interactions. In any of these situations, horizontal gene transfer is very difficult to measure, but is also a key evolutionary mechanism that maintains diversity and novel mutation.
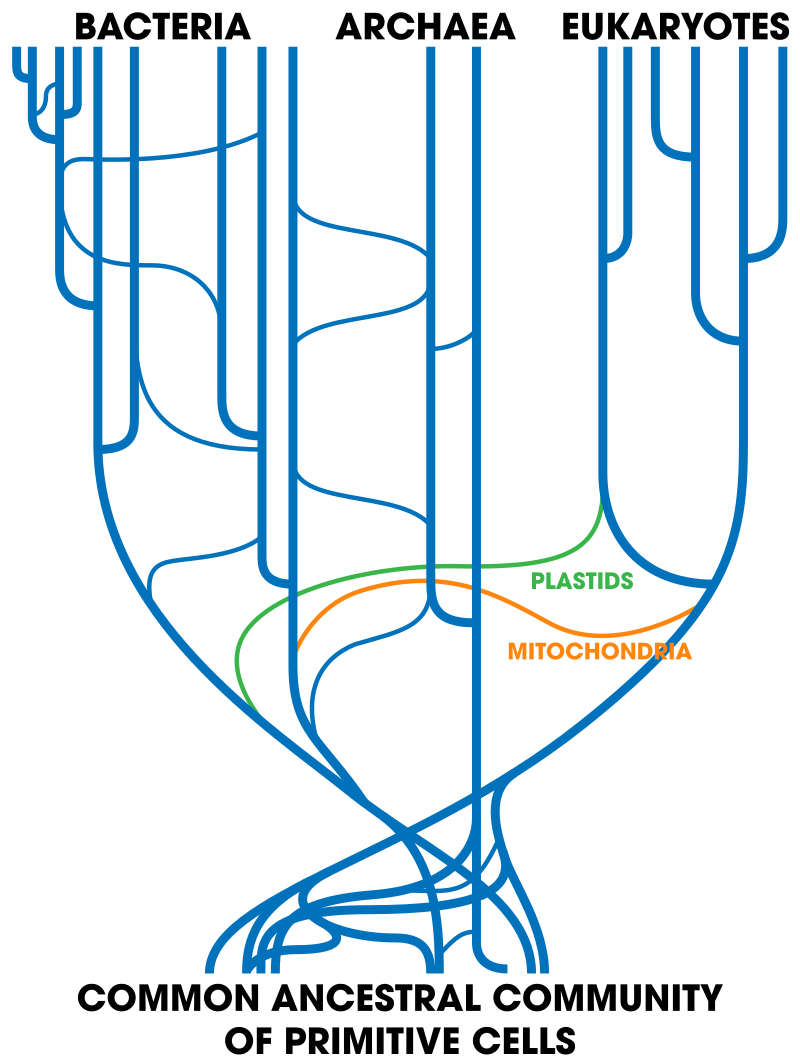
(Smets, 2005).
.svg){kind=link}
So far, all of the mechanisms of evolution discussed have been random in nature. Our final mechanism, natural selection, is very much not random. The genetic mutations produced by drift, horizontal migration, and others simply make an organism fit into its environment differently, which can lead to being passing such genes along or not. For natural selection to work, there are a number of important requirements that must be met. Within a population, there must be enough genetic variation to provide the flexibility necessary for advantageous mutations. There must also be a system of heredity that allow offspring to inherit the genes of the parents. Ultimately, a population will necessarily experience differential reproduction, where not all individuals will survive to reproduce due to environmental pressures. These can be everyday pressures, such as predation, that exist in periods of environmental calm, or stasis. They can also occur as a result of rapid environmental change, such as a local volcanic eruption, an extinction-triggering asteroid impact, or our modern episode of anthropogenic climate change. Natural Selection assures that the individuals who survive are more likely to pass on their genes.
Forms of Natural Selection
Some of the most amazing, flamboyant, and beautiful things in the biological world are the result of sexual selection. Sexual selection is where a species uses physical features or physical prowess to gain the opportunity to mate with another and pass on their genetic material. Peacock tails, colorful feathers on many male birds, and even dancing among fruit flies can inspire attraction of the opposite gender. Such things grab attention! Sexual selection can also produce situations that are very detrimental to a long life. Organism can become more susceptible to predation. Such selection can require the organism to give up its life in order to pass on these genes. Examples of the latter include sexual cannibalism in male black widow spiders or in some species of mantids where sperm is not released until the male’s head has been removed. In these mantids, this form of sexual selection may even be the main driver in the evolutionary change observed in their genitalia (Jensen et al., 2008).
Artificial Selection has been used for agriculture for thousands of years, as our species has continued to select for what are seen as favorable attributes. A classic example is the development of corn, or maize. Today, it is such a staple grain in our diet that you can find a corn product in nearly any food you purchase that has been processed. This includes meats, as many livestock are fed corn-based diets, even if they are not evolved to consume it well, such as with cattle. Native to Mesoamerica, the teosinte grass was used for its kernel and, over time, plants with plumper kernels were selected, eventually leading to the corn we see today. This form of plant domestication, an indigenous American export to the world, is now a staple everywhere.

(By John Doebley, Wikimedia Commons)
Another critical example of artificial selection, or domestication, that has shaped the modern world in particular came from Norman Borlaug’s work on wheat. Where ancient Mesoamericans were selecting for corn based simply upon the appearance of a kernel, Borlaug, as a microbiologist, would select wheat strains based upon their DNA. The problems he was addressing with wheat were directly related to a need to increase yield, shorten the stem of the grain to improve its ability to hold up until harvest, and to make it more disease resistant. Through the use of genetics, this work addressed these problems in the short span of 20 years. The results are still being experienced by people around the world today, through their ability to feed many more people than would have been possible otherwise (nobelprize.org, 2019). There are myriad examples of human-driven artificial selection that have resulted in the food we eat today. Artificial selection works by humans purposefully choosing individuals that will reproduce, based on which ones have characteristics valued by the farmer or researcher.
Coevolution
There are many examples of situations where two or more species will, as time passes, environments change, and natural selection occurs, affect one another’s evolutionary trajectory. Most often, these occur through ecological relationships. These relationships can be mutually beneficial, competitive, predatory, or parasitic. Coevolution is common in nature and illustrates the web of complicated relationships that allow the biosphere to function.
A classic example of coevolution is one that we rely on daily, the interaction between flowering plants and pollinators, such as honeybees. Such coevolutionary relationships are mutualistic, meaning that both species benefit from the interaction. The bees gain food while the plants are able to spread their pollen. Another excellent example of coevolution exists between the acacia ant (Pseudomyrmex ferruginea) and the acacia tree (Vachellia) that it protects. The ant is an obligate organism, in that this form of mutualism is absolutely necessary for its ability to reproduce. The acacia plants provide food and thorns that serve as nests while the ants protect the acacia from herbivores. Such examples are not limited to the world of the living. The fossil record even bears out examples. A fabulous one, depicted in the image below, shows a platycerid snail fossilized near the anal vent of a crinoid. There are actually quite a few examples of this particular relationship, which suggests that it was quite normal for these snails to feast upon crinoid poop!

(Rich Schrantz, 2000).
A Word About “Fitness”
Genetic change within populations acts randomly. Natural selection makes it more probable that individuals who are genetically prepared for current or new environmental realities will survive. They are the ones most likely to pass on the genes coded for the traits which permit survival. While genetic changes are random, natural selection is most definitely not random. Adaptations that make an organism fit for a cold climate, such as fur on a polar bear, will not likely be advantageous when that climate warms again at some point in the future. Staphylococcus aureus, a purveyor of infection throughout the human body, was famously stopped by the antibiotic Penicillin, developed by Alexander Fleming early in the last century. S. aureus roared back rapidly with resistance a few short years later and has now become so resistant to antibiotic treatment that some strains, such as “Methicillin-resistant S. aureus” (MRSA) are now feared in hospitals, where community infections can spread rapidly among healthy individuals (Chambers and DeLeo, 2010).
Evolution of a bacteria on a “Mega-Plate” petri dish, Koshony Lab, Harvard Medical School
Fitness is a situational circumstance, dependent upon being “good enough” for the environmental conditions of the moment, not the ability of an individual to “pull themselves up by their bootstraps”, or run a marathon, or win in a fight, providing themselves some kind of position of advantage through their own will. Ultimately the fitness of an individual is expressed by adaptations, or features that provide some improved function in an environment that is produced through natural selection. There are many features, it should be noted, that are not adaptations. These include vestigial structures, non-functional adaptations left over from an ancestor. Another example of a non-adaptive trait includes the red color of your own blood, which is a result of its chemistry and not due to some kind of selection. Exaptations make up another category. These are features that may have formerly had an adaptive purpose, but was not produced by natural selection for its current use. These can also be both physical and behavioral. One behavioral exaptation example might be domesticated dogs licking the mouths of their mothers. Interpreted as a means to get a parent to regurgitate food for them, this same behavior in wild wolves toward a lead wolf is interpreted as showing submissiveness (Bauer and Smuts, 2007).
Some traits can also be maladaptive, or harmful to a species fitness. A 2019 study released by Pagano et al. suggests that the eustachian tube construction in Neanderthal humans, Homo sapien neanderthalensis, made it susceptible to ear infections. Their reconstruction of the neanderthal eustachian tube suggests that its horizontal construction, very similar to human infants, may very well have even contributed to their eventual extinction. Human infants are notoriously susceptible to ear infections because of this construction of the tube, which can lead to pneumonia and, if left untreated, even death. As infants grow toward adolescence and adulthood, the eustachian tube does not remain horizontal, allowing drainage to occur and less likelihood of infections. Interestingly, while many of us (particularly of European descent) carry neanderthal genetic variants within our genome, given that our two sub-species are known to have interbred thousands of years ago, it is likely that this eustachian tube, arguably maladaptive in neanderthals, is a trait derived from a common ancestor. Ultimately, this trait may have reduced fitness for our nearest human cousins while remaining a relatively benign feature for our own species.
Modern Evolutionary Theory: “the Modern Synthesis”
The combination of Darwinian ideas with Mendel’s genetics is referred to as the “Modern Synthesis”. This term was coined by Julian Huxley in his 1942 book, “Evolution: The Modern Synthesis”. Tying in Malthusian ideas about population genetics with evolutionary theory, Huxley provided a framework that effectively brings together the macroevolutionary changes seen in the fossil record with the microevolutionary changes observed in nature and laboratories. However, this revolutionary model also provided the foundation for the rapid biological advances that would occur during the latter half of the 20th and into the 21st centuries. Some of these later developments and discoveries, such as horizontal gene transfer, developmental biology and embryology, etc. have yet to be fully integrated within this framework. Rather than totally replacing the modern synthesis with something new, work continues to discover ways to integrate such new fields and discoveries, ultimately towards finding a way to unite these ideas into a unified, single model. This can be referred to as neo-Darwinian evolutionary theory.

(Wikipedia Commons)
_____________
Chapter Contents
- 1 Evolution: Patterns in the Biosphere
- 1.1 Systematics: Organizing Life
- 1.2 Revolutionary Ideas: Darwin and Wallace
- 1.2.1 A New Paradigm Emerges: Darwin and Wallace
- 1.2.2 Meanwhile, Back in Europe: Mendelian Inheritance and Genetics Take Shape
- 1.2.3 Mutation, Migration, Drift, and Natural Selection
- 1.2.4 Forms of Natural Selection
- 1.2.5 Coevolution
- 1.2.6 A Word About “Fitness”
- 1.2.7 Modern Evolutionary Theory: “the Modern Synthesis”
- 1.3 Evolution: Making Sense of Patterns