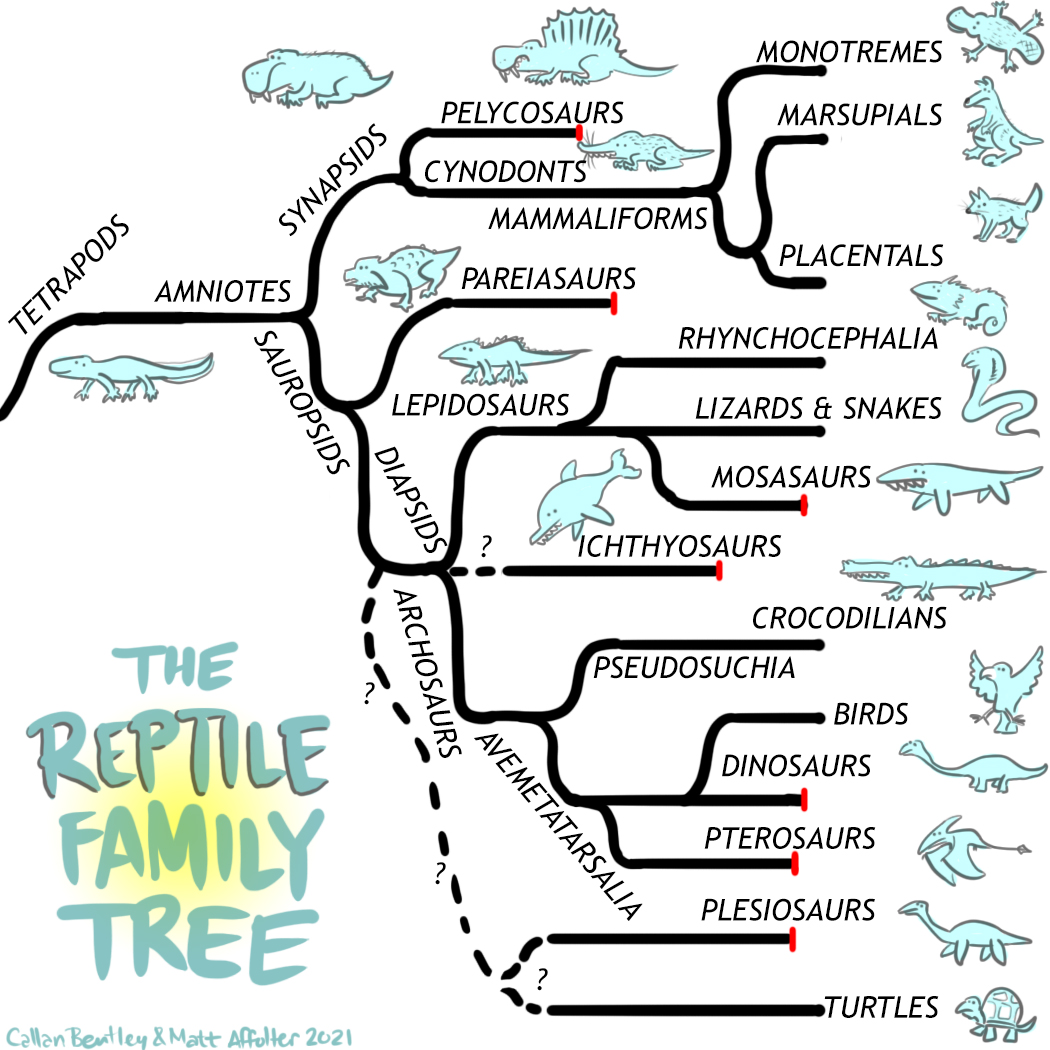
Reptile Radiation: Tetrapods Branch Out
After reading this chapter, students should be able to:
- Define what the term ‘reptile’ really means
- Discuss the early evolution of amniotes
- Understand the path from synapsids to modern mammals
- Interpret early sauropsids and their skull shapes
- Differentiate between important diapsids, like archosaurs, squamata, pterosaurs, and sauropterygia
Introduction

The term ‘reptile’ seems straightforward, but when discussing evolutionary history, the term becomes much more complex. Reptile, as a biologic term, is actually paraphyletic. This means that all reptiles are not in the same simple family tree, or, some members of the group are excluded. For example, consider birds. The term bird includes the first bird (such as Archaeopteryx or a related animal) and all of the descendants, including all living birds. However, birds evolved from dinosaurs, which are reptiles. Does that mean birds are reptiles? This is one reason why the word reptile is problematic. For some researchers, birds are reptiles. For others, they are not.

Another issue is where does the ‘reptile’ branch begin in the tree of life? To understand this, first one has to consider how reptiles evolved from non-reptile ancestors. The ancestors of the animals that eventually became reptiles were amphibians. During the mid-Paleozoic, the first fish to walk on land eventually turned into a common group of animals still alive today: amphibians. Amphibians were the first tetrapods (four-limbed animals) and they took advantage of the warm and tropical climate in the Devonian and Carboniferous and proliferated into many terrestrial niches with large and strange forms, like Eyrops.

Towards the end of the Paleozoic, as Pangea assembled and the climate dried out, amphibians were at a disadvantage. A new evolutionary leap occurred that allowed some animals to lay waterproof eggs on land, something their amphibian relatives were unable to do. This new group is the amniotes. Exactly which animal qualifies as the first amniote is a subject of debate, but Casineria or something similar is a good candidate. If this animal scurried in front of you right now, it probably would look like a modern reptile. However, it is actually not that closely related to modern lizards and it technically would not even be considered a true reptile.
Very early in the amniote lineage, a crucial divergence occurred that has a profound effect on our modern world. One branch became the synapsids, the group that would eventually become mammals. The other branch, called the sauropsids, contain the non-mammal tetrapods: modern reptiles, birds, dinosaurs, and a large array of extinct lineages. Sauropsid, therefore, is potentially the best synonym for the word reptile. However, some scientists exclude parareptiles, an early extinct clade that was common at the end of the Paleozoic, from the sauropsid lineage.
Synapsids and mammals

The synapsids, the group of tetrapods which would eventually lead to mammals and humans, started out with a very reptilian-like body plan. Synapsid means fused arch, referring to the hole near the base of the skull. This hole in the synapsid skull is correlated to the temple in your skull. The first group of synapsids to diversify and specialize were the pelycosaurs. The pelycosaurs were the dominant and largest group of tetrapods in the Permian, and were formerly known as the mammal-like reptiles. However, since they are not true mammals or reptiles, this name has fallen out of favor.

The most famous member of this group is Dimetrodon. Its name means ‘two-measure tooth’ because it had different teeth for different uses. This was one of the earliest animals found in the fossil record that have differentiated teeth for different purposes, which is a hallmark of mammals. It is more closely related to humans than dinosaurs. Though it is often portrayed with dinosaurs in kids’ books, it lived 40 million years before them. It lived in the Permian, and the first dinosaurs were Triassic. From track sites, it has been hypothesized that Dimetrodon could do a ‘high walk’ similar to what crocodiles can do, which is more efficient.
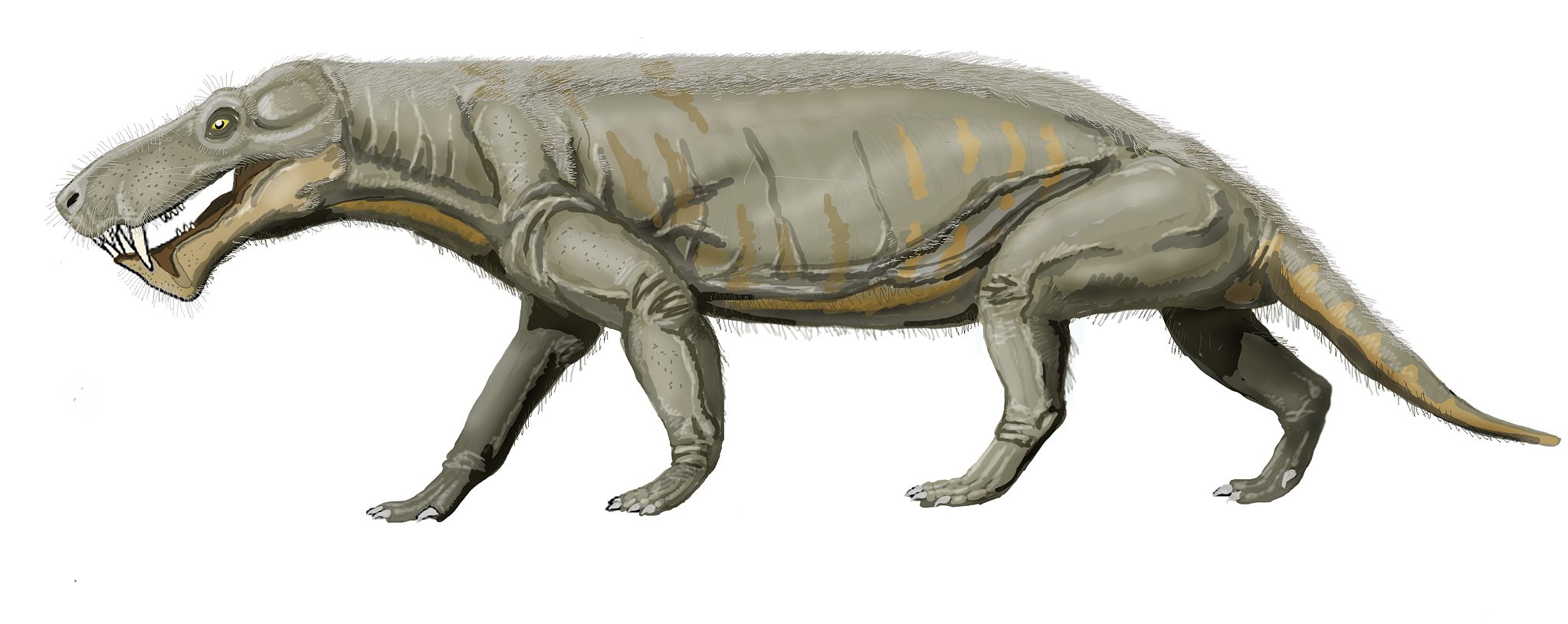
One of the branches of the pelycosaurs evolved into a new form called therapsids. These replaced pelycosaurs as the dominant land animals toward the end of the Permian. The therapsids had several advantages over their less derived pelycosaur relatives, features more akin to their modern mammal descendants. They had a fully-upright stance, with their limbs always beneath the body. There is also circumstantial evidence that they were endothermic (commonly known as warm-blooded) and had at least some hair. Scientists have found whisker pits in the skull, which is good evidence of hair. Examples of therapsids include the ferocious Gorgonops, one of the first saber-toothed animals.

Most of the therapsids went extinct at the end-Permian mass extinction, and even more went extinct at the end-Triassic mass extinction. One subgroup that survived both were the cynodonts. Meaning ‘dog tooth,’ cynodonts possessed even more mammalian traits.

An important but inconspicuous adaptation they evolved was a bony palate on the roof of their mouths. This has been attributed to a higher metabolism which matches its endothermy (warm-blooded), either by allowing breathing while chewing and/or allowing more complex chewing to help break down food.
True Mammals

By the mid Triassic, some cynodonts had evolved into true mammals. Technically, the first mammal is the last common ancestor of all living mammals, even if other animals (now extinct, called mammaliforms) had similar traits. Since many characteristic features of mammals (e.g. milk production (lactation), fur, neocortex of the brain) do not fossilize well, scientists commonly focus on jaw bones to identify and classify mammals.. Multiple bones found in the jaw of early synapsids migrated to form the bones of the inner ear. Since these ear bones are the smallest bones in mammals, they are also hard to find. However, the jaw is larger and more durable, so when the mandible fused into a single bone, it is a clear indication of a mammal or close ancestor. A common choice for the earliest animal to be considered a mammaliaform is Morganucodon.

The earliest mammals are most likely related to monotremes, whose living examples are limited to the platypus and echidna. Monotremes are mammals, yet they lay eggs and do not have nipples, and instead lactate by ‘sweating’ milk from their chest. They also have a single orifice used for urination, defecation, and reproduction, which is what the name ‘monotreme’ refers to. These more primitive features also support the evidence of monotremes being the first mammals. The details of the evolution of the mammal branches of the family tree, like marsupials (Metatheria) and placentals (Eutheria), is complicated. Comparing the genomes of the living groups put the divergence much further back than the fossil record does. Scientists can do this with molecular clock studies. By looking at the amount of mutations in related species, experts can estimate how long it has been since they evolutionarily diverged. Adding to the complexity is the fact that the earliest fossils of mammals are controversial as to their taxonomic placement. For example, the earliest proposed placental mammal (Juramaia) is Jurassic (Zou et al., 2011), and yet the earliest known monotreme (Teinolophus) is Cretaceous (Rich et al., 2001), even though placental mammals are more derived and should appear after monotremes.
Early Reptiles & Synapsids: Did I Get It?
Sauropsids
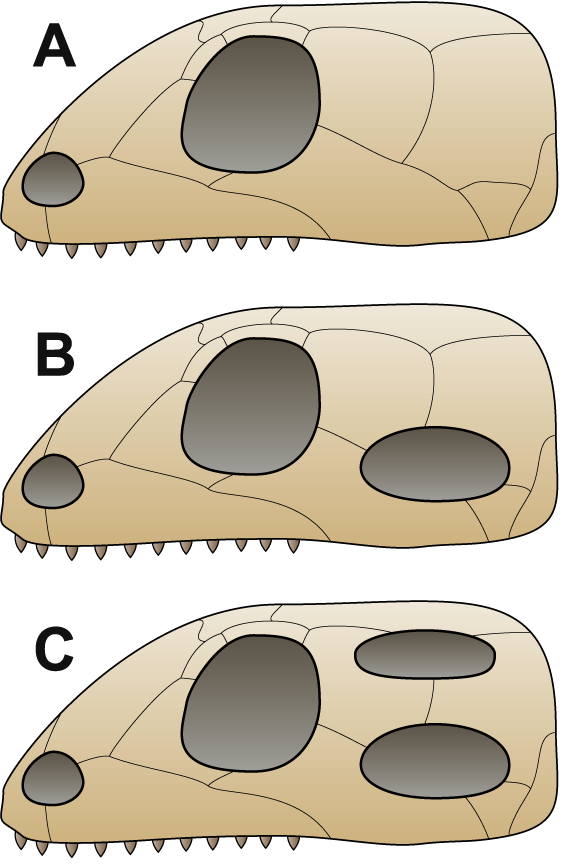
The amniotes had two major divisions in their early evolution: the synapsids, as discussed earlier, and the sauropsids, the branch that would eventually evolve into reptiles, dinosaurs, and birds. These early amniotes have been traditionally separated based on specific features of the skull. Two major groups make up the sauropsid branch: the anapsids and diapsids. Anapsids are reptile skulls with no openings in the rear of the skull, while diapsids have two such openings. As one might expect, the details of which of these three skull types was first and existed in the ancestral amniote (thus leading to the other types) is still controversial.

Anapsids are a clade that is roughly equivalent to the parareptiles, a group mentioned before in the section on early reptiles. Examples include the pariasaurs and the mesosaurs, the first tetrapod group to evolve an aquatic lifestyle since the tetrapods left the water in the first place. Mesosaurus fossils were also used by Alfred Wegener to reconstruct Pangea, since they lived in terrestrial aquatic habitats during the Permian. Recent analysis of anapsids have completely removed anapsids as a valid clade, with even mesosaurs showing skull openings and skull variability within the group. This means that traditional anapsid skulls may be more of a result of convergent evolution than inherited traits. Humans, for example, technically have an ‘anapsid’ skull since our only openings are our orbits. Regardless of where the exact positioning of parareptiles or mesosaurs is, it is inferred that this lineage went extinct. The only possible exception is turtles, which will be discussed later.
Diapsids

The other major branch of the sauropsid family tree is the diapsids. Their name means ‘double arch,’ referring to the two holes toward the back of the skull. Diapsids are true reptiles, and the theoretical most recent common ancestor of all living reptiles is almost certainly a diapsid. The earliest diapsids were the araeoscelidans, which resembled modern lizards except for their elongated limbs.

A sister group to the araeoscelidans branched into two major divisions: the lepidosaurs and archosaurs. The lepidosaurs (‘scaled lizards’) are famous for their overlapping scales. Within the lepidosaurs, Rhynchocephalia (’beak heads’) are interesting in that they were highly diverse and common across the planet in the Mesozoic. Today, however, only one species has survived: the tuatara, a lizard endemic to small islands off the coast of New Zealand.

Squamata is the other group within lepidosaurs. First evolving in the Triassic, Squamata contains nearly all modern lizards and snakes, which is the most diverse order of land vertebrates that exists on Earth. One of the more famous groups of animals that are extinct within Squamata are the mosasaurs. They were the apex predators of the Cretaceous seas and became the largest reptiles ever found in the oceans of Earth (at over 50 feet long), but went extinct with the non-avian dinosaurs. Their closest living relatives are snakes.
Archosaurs

Archosaurs are the other major division within the diapsids. Their evolutionary history is complicated and has many extinct groups of importance besides the dinosaurs. The name archosaur means ‘ruling lizard,’ and throughout the Mesozoic, they were the dominant land animals. As with other groups, the definition of what an archosaur is and what is a close relative depends on the use of the word. Traditionally, archosaur-like reptiles (e.g. not lepidosaur-like reptiles) could be considered archosaurs. However, the modern phylogenetic definition of an archosaur is the last common ancestor between crocodiles and birds. This means these archosaur-like reptiles are now considered archosauromorphs or archosauriforms. Which organisms were the first archosaurs is up for debate, but it may have been an animal like Protosaurus, a six-foot long lizard from the Permian.

In the Triassic, in the open landscape and ecospace following the Great Dying (AKA the end Permian Mass Extinction), the archosaurs diversified greatly. Many different lines of archosauromorphs that originated soon went extinct at the next extinction, the end-Triassic extinction. Interestingly, there was a large amount of convergent evolution with future forms that became more well known. An early example is Tanystropheus, which had a long neck like sauropod dinosaurs.
Why Triassic Animals Were Just the Weirdest

Another very strange and disputed early archosauromorph group are the drapanosaurs, which were tree-dwelling lizards that had a bird-like head, chameleon-like body, and a claw on the end of their prehensile tails.

Less in dispute, as far as being an early archosauromorph, are the rhynchosaurs. They had short, stocky bodies and were herbivores. They had the first complex chewing mechanism found on Earth, with rodent-like ‘incisors’ in the front of their mouths and large cheeks with densely-packed tooth batteries similar to a cow.

Within archosauriformes (the group more derived than archosauromorphs), but not true archosaurs, the most famous group would be Proterosuchidae. The crocodile-shaped animals had strange downward hooks on their upper jaws. Even though they are crocodile shaped, they are still only distantly related to modern crocodiles, and most likely looked similar due to them living in similar semi-aquatic habitats.
Pseudosuchia

Archosauria proper has two major divisions, nicknamed after the living animals that came from that group. Pseudosuchia, a name that means ‘false crocodiles,’ actually do contain modern crocodiles and therefore are called ‘crocodile-line archosaurs.’ Avemetatarsalia, which refers to the bird-like foot the animals have, are called ‘bird-line archosaurs.’ One important group which may be a sister group to both of these lines or may be on the Pseudosuchia line are the phytosaurs. They were also crocodilian in shape, but had noses in the center of their skull instead of at the end of the snout. Despite their name (‘plant lizards’), they were actually apex predators in the Triassic.
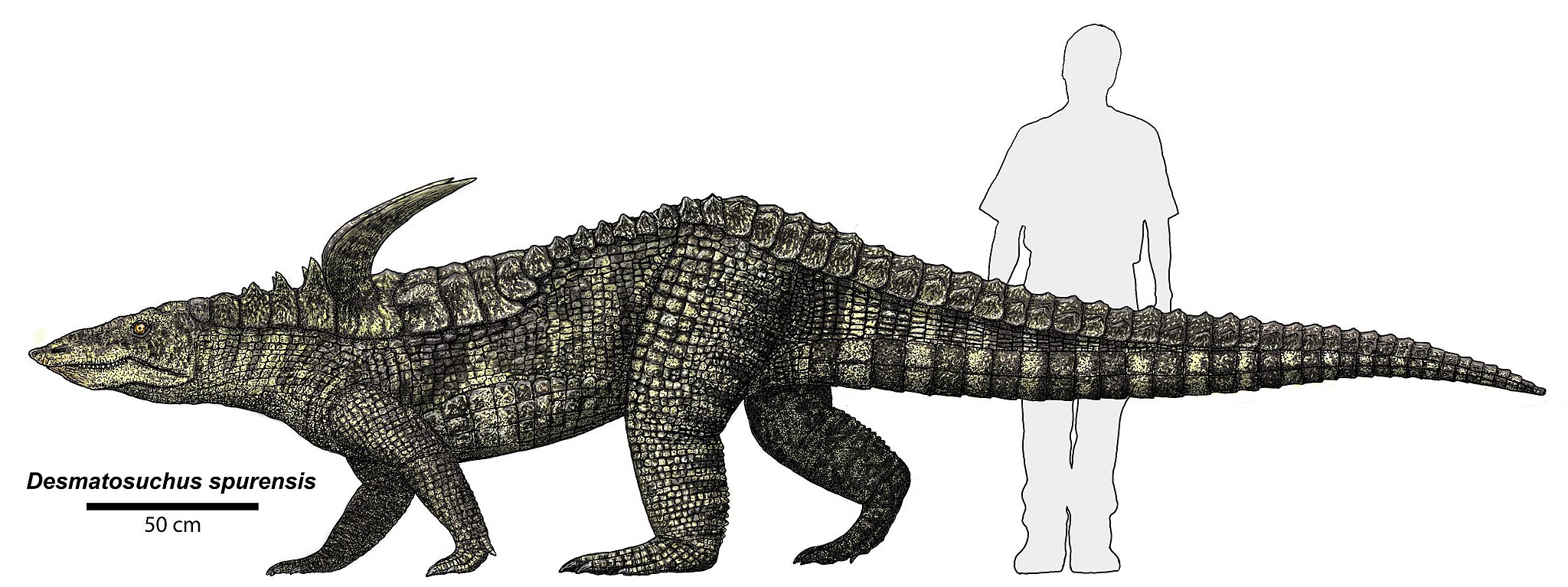
Within Pseudosuchia, there are many different groups, but one of the more interesting are the aetosaurs, another classic example of convergent evolution. They had snouts like pigs which aided in their terrestrial herbivore lifestyle. They were also covered in armor, superficially similar to the ankylosaurs which would come along more than 50 million years later.

Actual crocodiles, or at least the group Crocodylomorpha, first appeared in the late Triassic. Even within this group, there is great diversity of forms and lifestyles. This includes fully aquatic crocs, fully terrestrial crocs, crocs with hooves, armored crocs, and even vegetarian crocs. Many of these more unusual forms went extinct with the dinosaurs, leaving the crocs we know today. The first modern crocodilians appeared toward the end of the Cretaceous.
Sauropsids, Diapsids, Archosaurs, and Pseudosuchia: Did I Get It?
Avemetatarsalia
Pterosaurs
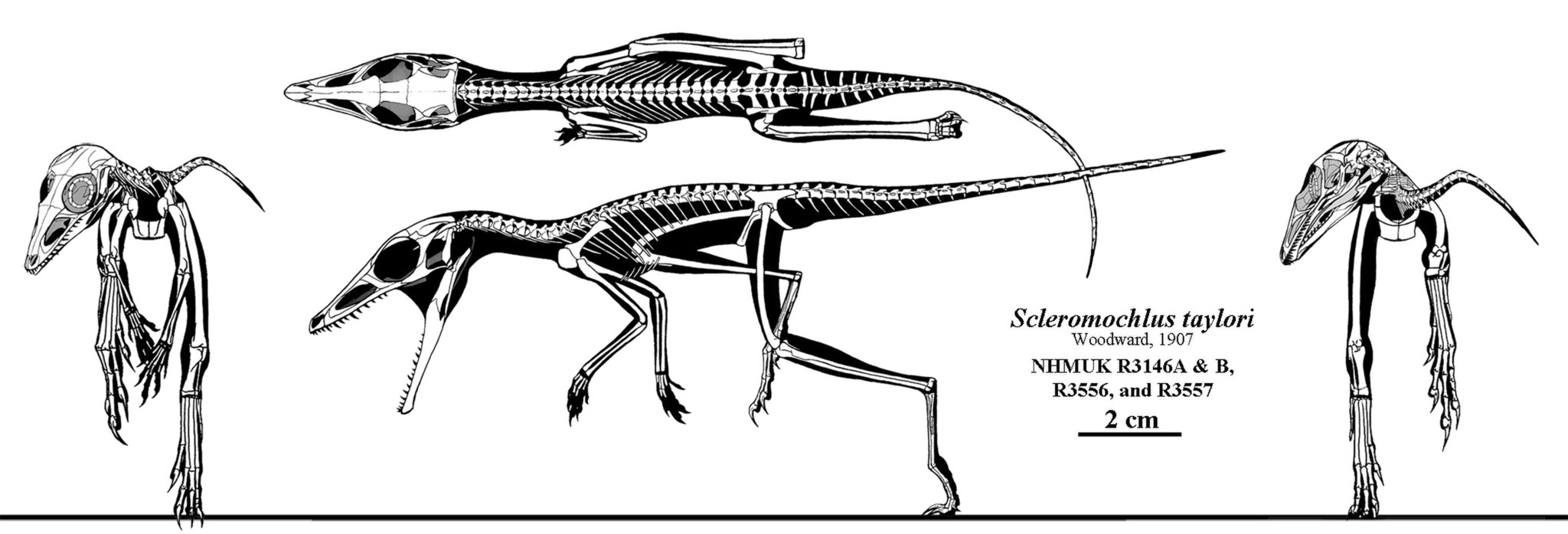
Within the bird-line archosaurs, Avemetatarsalia, the most famous early group is the pterosaurs. In fact, a similar term that is used is almost as a synonym for Avemetatarsalia is Ornithodira, which is defined as the clade that contains the most recent ancestor of all dinosaurs and pterosaurs. These flying reptiles (the first vertebrates ever to evolve powered flight) have an even more complex and mysterious evolutionary history than other groups due to the thin, fragile nature of their skeletons. One possible ancestor to the pterosaurs is Scleromochlus, a Triassic animal that spent its time hopping. If this animal is not an early pterosaur, it may be an early avemetatarsalian, or even an early archosauramorph.

All pterosaurs had a long fifth finger that extended to form their wing. The pterosaurs have two major groups within their clade. The rhamphorhynchoids had toothed beaks and long tails. Though fish are traditionally seen to be the main diet of these flyers, some had sharp teeth for tearing terrestrial animal flesh. The namesake of the group, Rhamphorhynchus, also had a rudder-like tail.
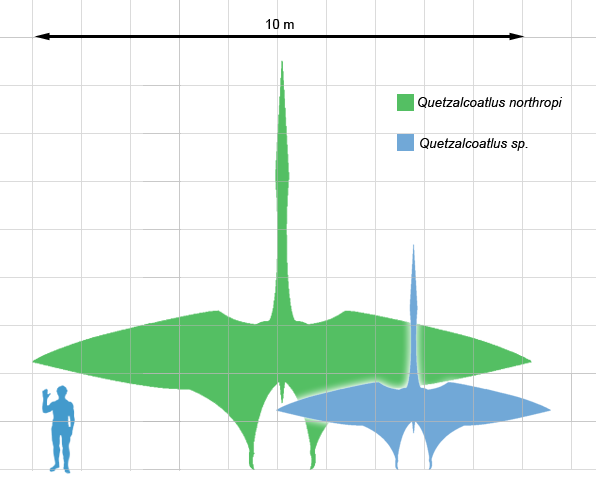
The other group of pterosaurs are the pterodactyloids. This name derives from Pterodactylus, a famous member of the pterodactyloids. As an aside, the term ‘pterodactyl’ is commonly used in place of pterosaur to refer to all flying reptiles, though that is incorrect. It should only refer to Pterodactylus and nothing else. Pteranodon, another famous pterodactyloid, gives the name ‘pterosaur’ to the overall group. Pterodactyloids are known for a lack of tails and more elaborate crests on their skulls. The most remarkable member of this family is Quetzalcoatlus. Named after the Aztec feathered serpent god, it is the largest animal ever known to fly at a wingspan of 36 feet!
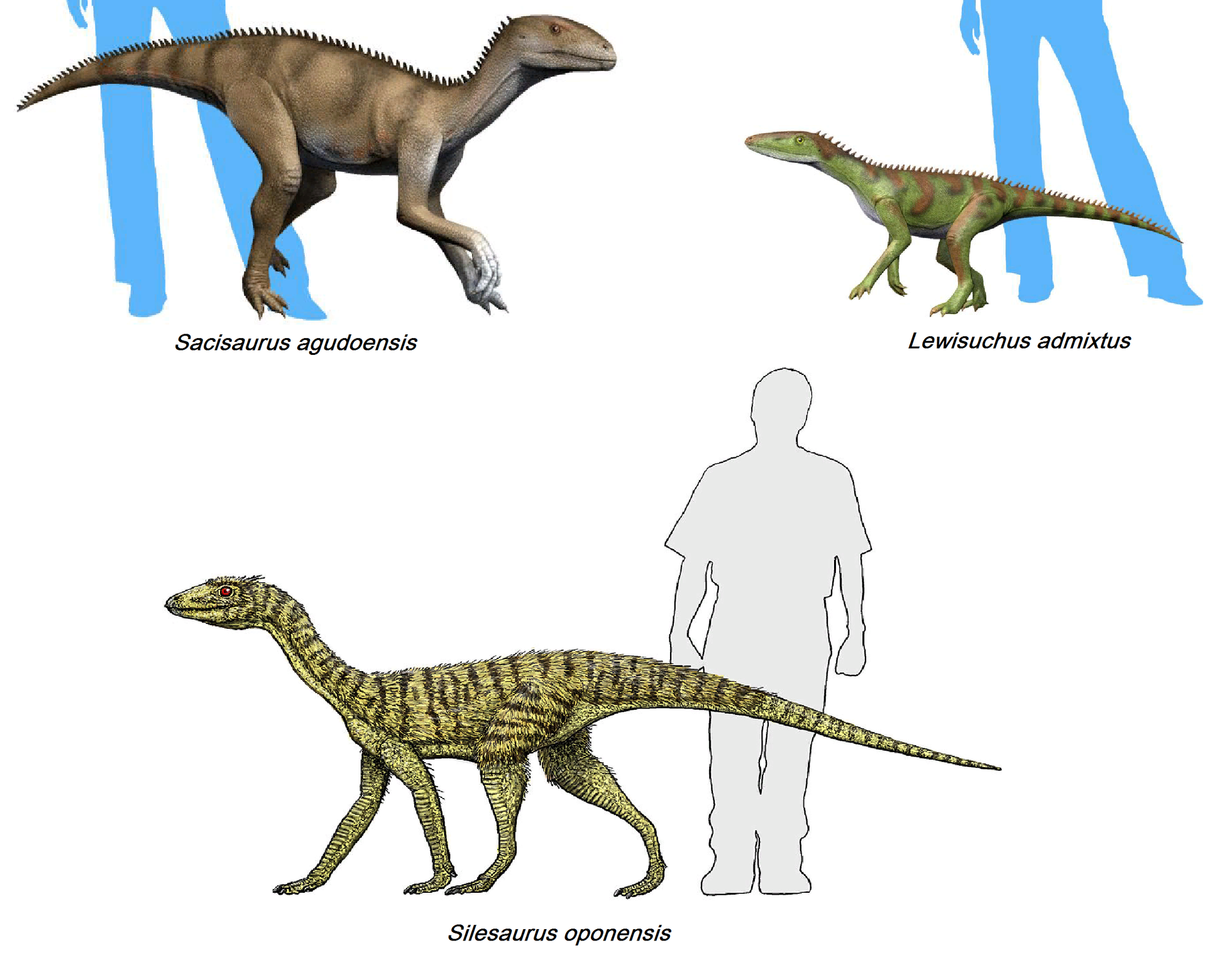
The more evolved bird-line archosaurs are generally called dinosauromorphs. The animals that are dinosauromorphs but not dinosaurs are generally small carnivores. To a novice, they would certainly look like dinosaurs. The group that is closest to dinosaurs are the silesaurs. They are currently classified as a sister group to Dinosauria. For more details on dinosaurs, go to Dinosaur Classification.
Ichthyosaurs

One group that has an uncertain placement within the diapsid lineage (but are still generally considered diapsids) are the ichthyosaurs. These marine reptiles were an early branch on the archosaur tree, because they were already prolific in the early Triassic. Part of the difficulty in their placement phylogenetically comes from the lack of transitional fossils from land-dwelling ancestors to fully aquatic ichthyosaurs. Because of this uncertainty, various paleontologists have placed them as basal diapsids, as lepidosaurs, or as archosaurs. There is no clear consensus.

Ichthyosaurs, much like the creatures whom they convergently share a body plan – sharks and dolphins – were fast swimmers that hunted fish and other ocean animals, like ammonites. They even gave live birth, with a remarkable fossil preserving this process. They were some of the top predators in the ocean and filled many niches. For example, many had large eyes, suggesting they dove to great depths to hunt. Even with all their success, by the mid-Cretaceous, they were largely extinct, possibly out-competed by the mosasaurs.
Turtles and Sauropterygia
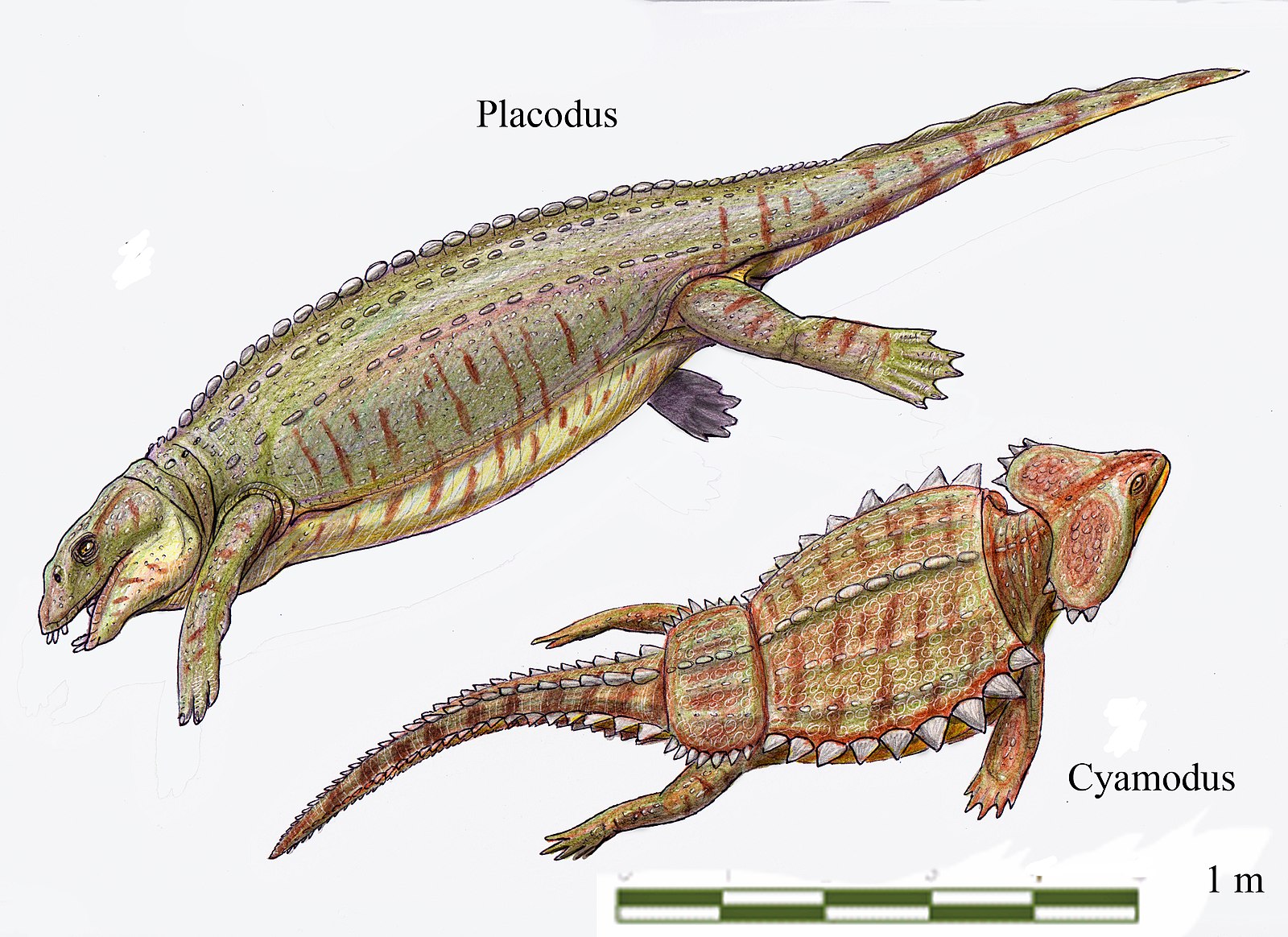
There is one other major marine reptile group and one other major living reptile group that has not been covered yet in this case study, and it turns out they are related. These groups are plesiosaurs and turtles. However different these groups may seem, the current established phylogeny has them related. One early (Triassic) group that shows some crossover between the two were the placodonts. They were marine with tough jaws and body armor, in some cases being full shells.

The group that includes the placodonts and plesiosaurs is called Sauroptergia, which means ‘lizard flipper.’ Along with the plesiosaurs, other groups like the nothosaurs and pachypleurosaurs lived along side the placodonts in the Triassic. At the end of the Triassic, all of the groups went extinct except for the branch of this group became the plesiosaurs, the only members of their superorder to survive the Triassic mass extinction. In the Jurassic and Cretaceous, they became very diverse and widespread in the oceans, alongside the ichthyosaurs (mostly earlier) and mosasaurs (mostly later). Their body plan featured a long neck and small head for grabbing fish, except for the subgroup known as pliosaurs, which had larger jaws and shorter necks. Like the other large marine reptiles in the Mesozoic, they went extinct at the same time as the dinosaurs 66 million years ago. They are probably most famous today for being the supposed creature known as the Loch Ness Monster. There are many reasons why to doubt this, including early evidence that turned out to be a hoax, but scientists have even attacked the specific reasons why it could not be a plesiosaur.
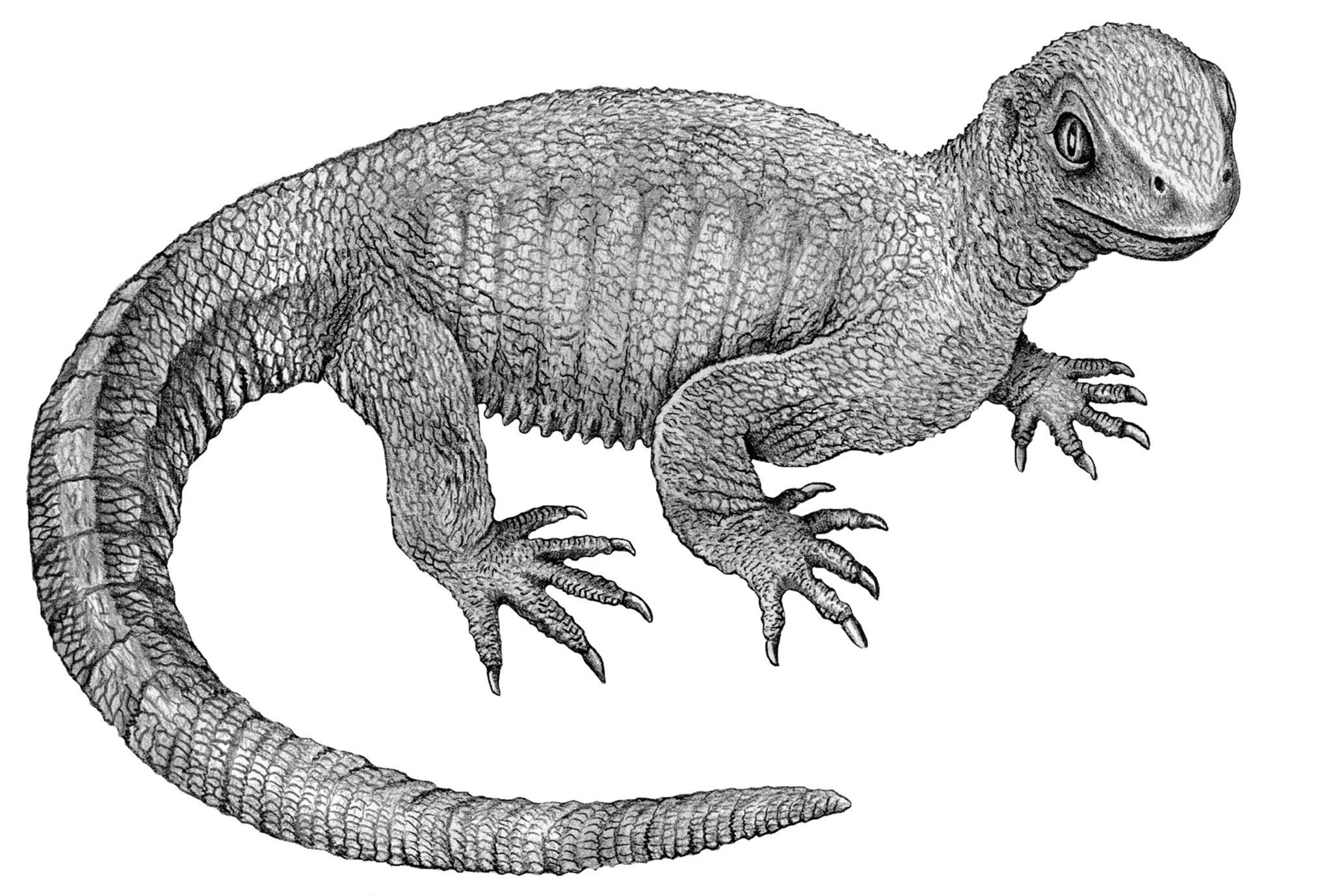
Knowing where turtles fit into the tetrapod family tree can help with the placement of Sauropterygia, but that is no easy task. Traditionally, turtles were considered anapsids. This comes from the fact that turtles show no hole openings in their fortified skulls. Also, turtles excrete urine more similarly to mammals, potentially pointing to a very basal position in their split from other reptiles, which would match anapsids well. However, early turtles like Pappochelys have been shown to be diapsids. They started out as land animals, potentially using their reinforced torsos to help with digging. Millions of years passed before they diversified into the multitude of forms seen today, including venturing back into the sea as the largest living marine reptiles. Genetic evidence, comparing them to living archosaurs and lepidosaurs generally favors them as more closely related to archosaurs, but the debate certainly is still open. Some genetic clock studies place the divergence between turtles and other tetrapods all the way back in the mid Carboniferous, adding uncertainty to how primitive turtles might be.
Avemetatarsalia, Ichthyosaurs, and Sauropterygia: Did I Get It?
Final Thoughts
The evolution of tetrapods may be very complex and controversial, but understanding this process can not only give you a great understanding of the animals that inhabit our planet today, but also beautifully demonstrate many of the founding principles of evolution. While dinosaurs are the group that gets the most attention, there was a whole array of interesting reptiles that are not dinosaurs. Some of these non-dinosaur reptiles share similar features with dinosaurs because of their common amniote tetrapod ancestors–they all have four limbs and certain features of the skull. There are also many wonderful examples of analogous traits via convergent evolution in the reptilian family tree due to them living in similar ecological niches (e.g. pliosaurs and crocodilians). The diversification of reptiles is an extraordinary example of adaptive radiation. An adaptive radiation is when a group of organisms diversifies to fill all potential ecological niches to fully take advantage of all available resources. The expansion of reptiles to land, sea, and air exemplifies why the Mesozoic was historically dubbed “The Age of Reptiles.”
Further Reading
Rich, T. H., et al. (2001) “Monotreme nature of the Australian Early Cretaceous mammal Teinolophos”. Acta Palaeontologica Polonica 46(1). Pages 113–118.
“Your Inner Reptile,” PBS television program (2014): https://www.pbs.org/video/your-inner-fish-program-your-inner-reptile/
Zhe-Xi Luo; Chong-Xi Yuan; Qing-Jin Meng; Qiang Ji (2011). “A Jurassic eutherian mammal and divergence of marsupials and placentals”
Chapter Contents