
Image: Wikipedia, Billwhittaker, CC BY-SA 3.0
STUDENT LEARNING OUTCOMES
At the end of this chapter, students should be able to:
- Discuss Homo sapiens sapiens in the larger context of hominid evolution;
- Highlight some of the critical splits in the human phylogenetic tree;
- Discuss some of the features of hominid skull anatomy that can be used for comparative genotype analysis between hominid groups;
- Describe how humans migrated from Africa, including the timing and pathways;
- Describe how we can trace these migrations using paternal and maternal haplogroups;
- Identify some of the key technological innovations during the evolution of hominids, particularly among anatomically modern humans and Neanderthals;
- Understand how human evolution is very similar to evolution within other taxonomic families of organisms.
Humans as a species
The evolution of humans began 13.8 billion years ago with the Big Bang itself, which set in motion the forces and energy that would make our history all possible (though, not inevitable!). Elements created and compounds formed after this event would eventually form our planet, Earth. At some point after 4 billion years ago, living, self-replicating materials formed that would become our first ancestors. In order for this to happen, these early life forms had to use resources from their environment to survive and thrive.
So it is with humans today. All life extracts energy and nutrients from its environment for respiration, digestion, and procreation. Our species, and some of our ancestors, took this extraction to a new level. They developed tools. These tools made resource extraction much more effective, leading to new evolutionary pressures, more adaptations, and eventually, you and me.
This case study examines the development of our species. The story is a long one, so this is merely a summary. Ultimately, it serves to highlight several aspects of humans. Two threads of hominid evolution are emphasized. We explore the evolution of tool use among hominids as a way of exploring the development of human intelligence and our species’ ever-increasing ability to extract resources from its environment. We also explore some of the particular anatomical features of hominid skulls as a way of showcasing hominid physiological development.
Here are some of the key threads that are woven together in this chapter:
- Like other organisms, anatomically modern humans evolved in a manner consistent with evolutionary theory.
- Hominid evolution can be traced using fossils and comparative anatomy.
- Like other organisms, humans extract energy and alter their environment to survive.
- Human ingenuity with tools began deep in hominid history, and is not unique to anatomically modern humans.
Before you dive into the specifically human part of the story, you are encouraged to begin with a larger view of our mammalian past as a review. Watch the video below:
Paleoanthropology
If you read about human evolution and the field of paleoanthropology, you will also encounter other primates you may consider old friends (e.g., chimpanzees) and a bunch of unusual primates who are extinct. You will also explore elements of the field of archaeology, the study of material culture. This last piece is pretty unique to research into hominid evolution, as there is no archaeological record for other organisms. The closest analog we get for archaeological materials in the rest of the living world is ichnology, the study of trace fossils. Archaeology is divided into two subfields. Historical archaeology explores human culture after the development of writing. Prehistorical archaeology explores everything else. Archaeology is typically a subfield of anthropology, the study of humans.
Paleoanthropology is a confluence of paleontology and anthropology that seeks to understand our human ancestry and associated culture. The field seeks a holistic exploration. This means that paleoanthropologists are interested in human biology, social structure, cultural development, material culture (archaeology), linguistic development, and more.
There have been many rather famous paleoanthropologists. This is because the field tends to engender a good deal of interest among people outside of the scientific community. Famous names include many members of the Leakey family. Louis and Mary Leakey and their son Richard Leakey who, with his partner Meave Leakey, have contributed much of what we know about our shared African ancestry. Their work in locations like Olduvai Gorge, Tanzania and nearby environs has provided important windows into how humans evolved, migrated, and are related to our primate cousins. Ian Tattersall, a curator at the American Museum of Natural History in New York City, has worked to understand more deeply our connection to current primate cousins but also to standardize terminology in the field. Still others, like Sonia Harmand, have dedicated their research to studying human cultural development, as exemplified in her work with stone tools.
Paleoanthropology is a dynamic and very young field. New insights into our human past are being published almost monthly. To keep up with these changes, refer to two particular locations included in the section on further reading at the end, the Smithsonian Institution’s human evolution program and the Institute of Human Origins at Arizona State University.
Hominid phylogeny terminology
As you dive into the study of human origins, you will encounter a bunch of confusing terms. We will begin by outlining some of these here.
Primate – Primates are a large group of social mammals that arose between 85 Ma and 55 Ma, probably from small tree-dwelling mammals. Shared characteristics of primates include relatively large brains, visual acuity, color vision, dexterous hands (prehensile with opposable thumbs for grasping), and large ranges of motion in their shoulder joints. Primates are a part of the order Primata, which contains two suborders, strepsirrhines and haplorhines. Humans are a member of this latter suborder.
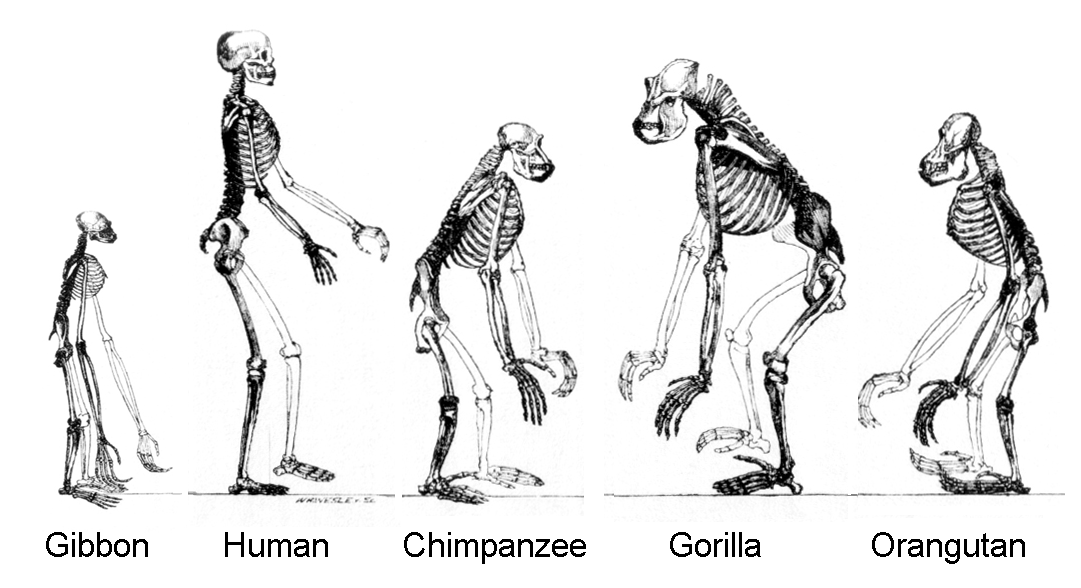
Image: Wikimedia, Public Domain
Hominidae – The hominids are the human taxonomic family. It is more colloquially referred to as the “great apes”. The term “hominid” is sometimes also used generally. Within this family, there are currently eight extant (living) species, contained within four genera. These species include us, Homo sapiens, Orangutans (Pongo abelii, Pongo tapanuliensis, Pongo pygmaeus), Gorillas (Gorilla gorilla, Gorilla beringei), and Chimpanzees (Pan troglodytes, Pan paniscus).
Homininae – The hominins are our subfamily and refers specifically to “African Apes”. This subfamily is divided into two “tribes” (the term is no connection to that which is used for indigenous human groups), Hominini and Gorillini.
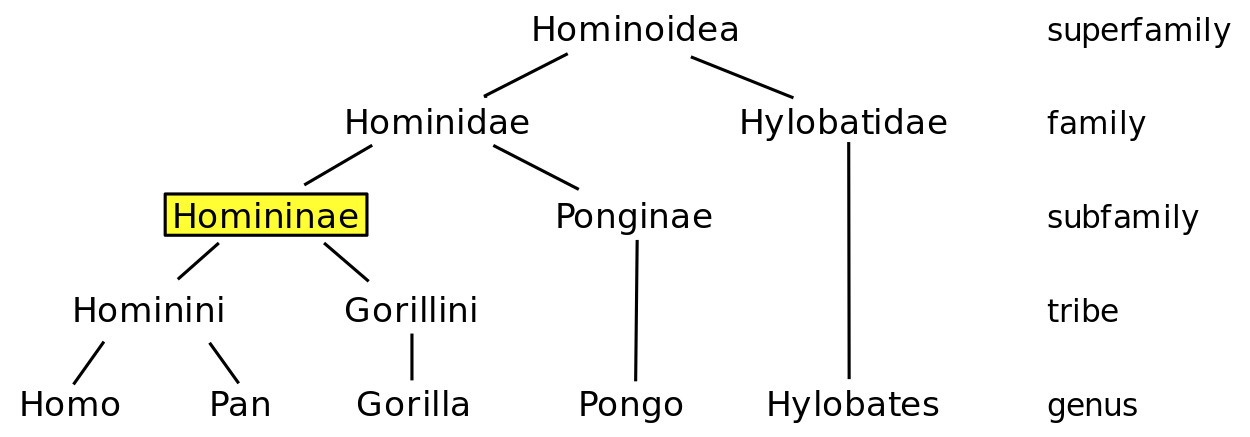
(Image: Wikimedia, By EoD – Own work, CC BY-SA 3.0)
Hominini – The hominini is a taxonomic tribe that includes our human ancestors and chimpanzees – two genera only. Sometimes chimps (Pan sp.) are separated out for their own tribe. Ultimately, the search for a common human ancestor between our close cousins chimpanzees extends to about 4-8 Ma (Patterson et al., 2006) but the split probably happened around 6 Ma. Once this split occurs, the phylogenetic tree is entirely focused on our ancestral lineage, all of whom are now extinct except for us. These genera include Homo and Australopithecus.
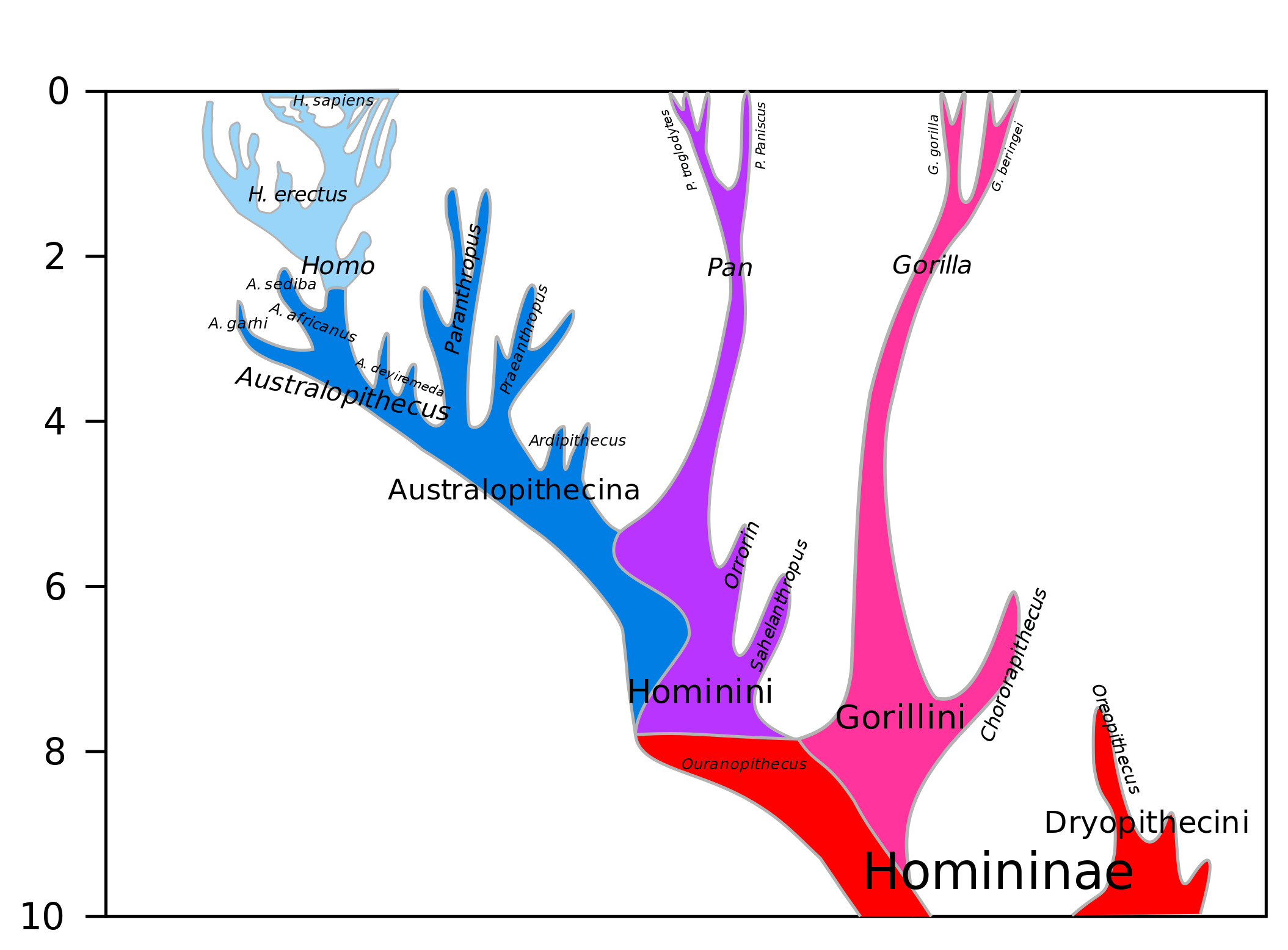
Image: Wikimedia, Dbachmann – Own work, CC BY-SA 4.0
Did I Get It? Who are Hominids?
Hominid Skull Terminology
Below there will be a good deal of discussion that uses hominid skull development to discuss hominid evolutionary change. Certainly, there are other anatomical features to use, when there are more complete skeletal remains. Skulls and skull fragments are probably the most common single fossil, so they are used in this case study. Below is a summary of some of the major parts of hominid skull anatomy.
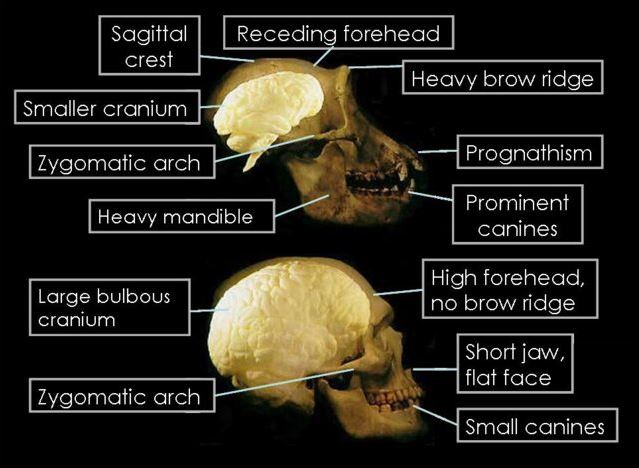
Image: Public Domain
If we look in detail at parts of the skull, some of the major areas become important in the story of human development. The first of these is the mandible, or lower jaw. A key difference between apes and humans in this feature is sheer size. Apes eat a great deal more tough, fibrous material than humans do, the chewing of which requires a more robust lower jaw. Humans also developed an accentuated chin structure to help provide some additional strength to what is a much weaker area of our lower jaw.
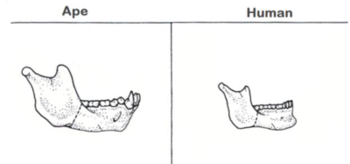
Image: Public Domain
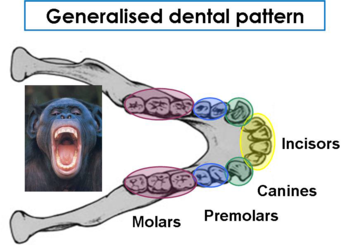
Image: Public Domain
The arrangement of teeth in apes and hominids is very similar. The dental arrangement includes molars and premolars for chewing, incisors for cutting, and canine teeth for tearing. Teeth are also one way that apes, as opposed to humans, can display sexual dimorphism (differences between genders). Male apes tend to have much larger canine teeth than female apes. There is no real advantage to the larger canine except for sexual selection, with females giving preference over one male for this particular feature over another. While certainly humans do notice one another’s teeth, mates are not selected based upon the size of their canine teeth. One other major difference between ape and human teeth is enamel thickness, with human tooth enamel being much thicker.
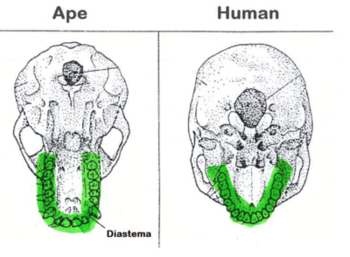
Image: Public Domain
On the upper jaw, the dental arcade provides additional areas where we can look for physiological changes in hominids. The shape of the dental arcade is an obvious starting point, where humans have a much more V-shaped arrangement and apes a more U-shaped arrangement. This is largely due to the shortening and flattening of the face through evolutionary change.
The other large difference here is the diastema, or the gap between the premolars and incisors where the large canine teeth reside. Humans have no diastema while hominid ancestors do.
Three other skull features are related to chewing, or lower jaw motion. These are the zygomaticus arch, the sagittal crest, and the supraorbital brow ridge.
The sagittal crest was lost to hominids as the demands of chewing became less strenuous. Because these crests were a primary area of attachment for the temporalis muscles that control the lower jaw, they could be quite pronounced in very early hominids.
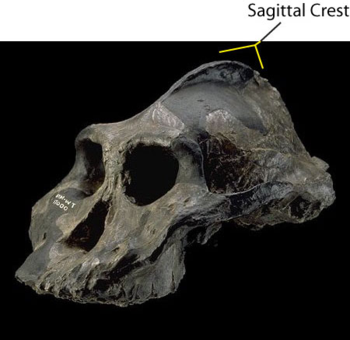
Image: Public Domain
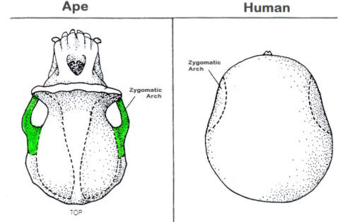
Image: Public Domain
The zygomaticus arch is much larger in earlier primates than it is in humans. This is due to the need for much thicker and robust temporal muscles. These muscles attached to the lower jaw through and behind the zygomaticus arch and then attach to the side of the cranium. They are the muscles that move the lower jaw and, the larger they are, the more powerful the chewing ability of the organism.
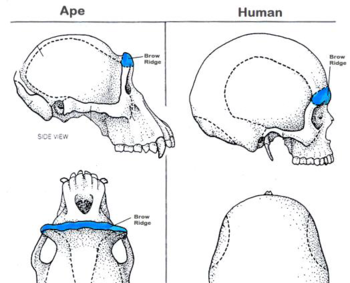
Image: Public Domain
The supraorbital brow ridge is one of the last skull features lost in hominid evolution, with humans no longer displaying it at all. This structure provided strength to the facial portion of the skull. As the powerful temporal muscles placed a good deal of strain on the face as they pulled on the cranium while food was being masticated. This ridge kept the eye sockets from collapsing under this stress.
Other features provide support for changes in locomotion, specifically the evolution from walking on all fours to mostly on the hind legs to full bipedalism, or walking upright on two feet. Today, humans are bipedal. Skull features that accompany this progressive change in stance include the nuchal area and the foramen magnum.
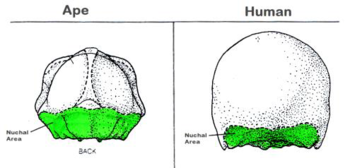
Image: Public Domain
The nuchal (pronounced “new-kul”) area is located on the base toward the rear of the skull. This is the area where neck muscles attach to the skull. Our primate ancestors have a much larger nuchal area. This is because much larger muscles are required to keep the individual looking forward when the spine is situated so far back in the skull.
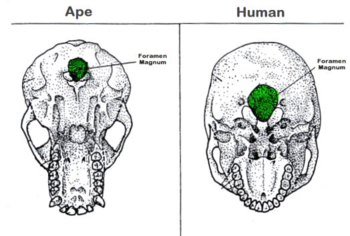
Image: Public Domain
The foramen magnum is the hole in the base of the skull where the spinal cord enters. Depending upon the primate species, the foramen magnum is located at a fulcrum, or balance point, and so its position is highly indicative of overall locomotion. In humans, it is in a more forward position while in apes, who walk mostly on all fours, it is situated further toward the rear of the skull.
The final and perhaps most peculiar change in the hominid skull is the cranial capacity. Until the genus Homo sp. evolves, hominid skull capacity does not change in any substantial way. However, it certainly increases quite a bit in our most recent ancestors. In Homo habilis, which will be discussed below, the cranial volume is a mere 600 cm3. In modern humans, by contrast, it is 1400 cm3. Some Neanderthals even exceeded this, with cranial volumes approaching 1700 cm3.
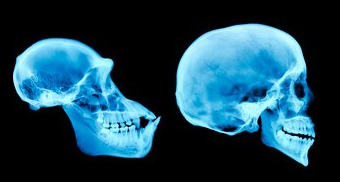
Image: Public Domain
The greater cranial volume evolved to fit a growing brain. One might be tempted to attribute this larger brain simply to greater intelligence, but this is a matter of great debate. While there does seem to be a correlation between the two variables, it is not necessarily causal, as intelligence (however you define that term) can potentially be attributed to other characteristics of the brain. Further, larger cranial capacities are also directly associated with larger body size, and adaptations to colder climates (which certainly was necessary in Neanderthals and modern humans). There is also some degree of sexual dimorphism among hominids. On average, males tend to have slightly larger cranial capacities than females of various hominid species. This is not an issue of greater intelligence, but rather cranial capacity being directly linked to larger average body size.
This is not to say that intelligence does not play a role in cranial capacity. It is more likely that brain physiology controls that. For decades, it was assumed that brain growth in humans was linked to a kind of reinforcing feedback loop between intelligence, technological development, and hunting meat. There may be some truth to this and, in the context of a discussion of Homo habilis below, the connection between meat and brain growth will be explored more specifically. Overall, as hominids evolved from a strictly vegetarian diet to scavenging bone marrow to directly hunting game, cranial capacity and brain growth followed. These were accentuated by technological developments that made scavenging and hunting more efficient. Simple wooden tools gave way to knapped stone scrapers. These gave way to elaborate arrow and spear points. Eventually – humans learned to smelt metals. Copper was followed by bronze and iron, each with their own “age,” and now there’s silicon. Are these all the result of greater cranial capacity and brain size? Perhaps some of it is, but certainly there are many other variables to consider.
Below is an excellent video showcasing hominid skull features by Dr. Daniel Lieberman, Professor of Human Evolutionary Biology at Harvard University.
Did I Get It? Hominid Skull Physiology
Highlights of our family tree: Australopithecina and Homo
Like all families, the human family and its ancestry are filled with interesting, odd, characters with unique traits and, in this case, a trail of important evolutionary changes and adaptations. The image below can be referred back to for context as we move through the human ancestral lineage, in this case, from right to left (note the vertical time scale). The image below does not include the first four hominids discussed below, all of which predate Australopithecus anamensis.
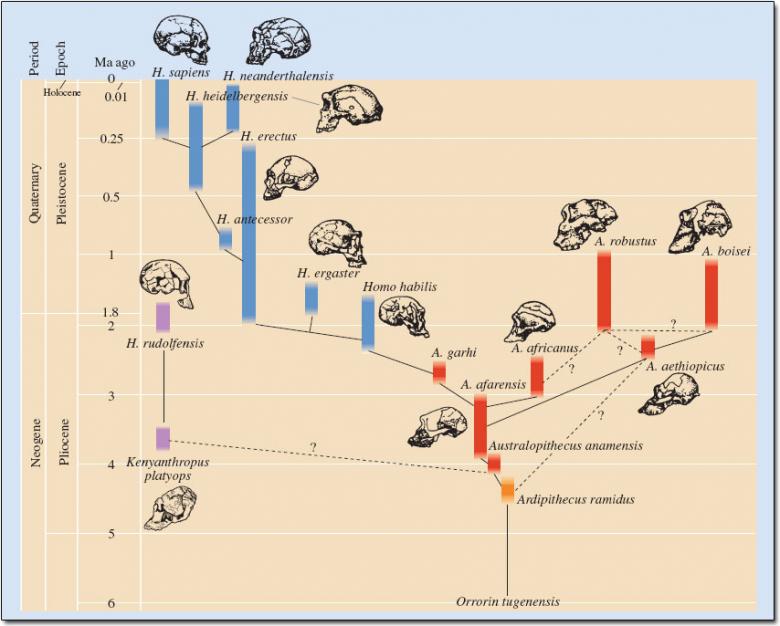
Image: Public Domain
Sahelanthropus tchadensis – 6-7 Ma

Image: Smithsonian Institution
Our oldest directly connected human ancestor, Sahelanthropus tchadensis, is traced to the modern African nation of Chad about 6-7 Ma. Fossil evidence suggests that they walked upright. The main evidence for this is the more forward positioning of the foramen magnum, the opening for the spinal cord at the base of the skull. As this opening evolutionarily moves forward, it would indicate that the spinal column was moving to a position directly below the skull, as it is for modern humans today. Sahelanthropous tchadensis is known only from nine cranial specimens (the first of which was discovered by Michael Brunet in 2001), so fossil evidence for much else is very limited. We can also say that, due to its heavy tooth wear, it was likely a vegetarian.
Skull fragment of Sahelanthropus tchadensis. Note the human-like feature in the downward and forward positioned foramen magnum and the ape-like protruding mouth, large teeth, narrow and small braincase, and highly pronounced occipital ridge above the eyes. (Model: Smithsonian Institution, Creative Commons License)
Orrorin tugenensis and Ardipithecus kadabba – 5-6 Ma
Little is known about these two hominids. Both come from east Africa: Kenya and Ethiopia, respectively. From the few skull and leg fragments known, there are two characteristics that they share with modern humans. There is one upper femur specimen of Orrorin tugenensis that shows the same kind of bone buildup typical of a biped. Dentition, or teeth, are also important for both of these hominids. Both had thick enamel, which is a shared characteristic with modern humans. Both were about the size of a modern chimpanzee and likely subsisted on a vegetarian diet.
Ardipithecus ramidus – 4.4 Ma
Known as “Ardi” the first partial skeleton of this organism was presented in 2009, though Ardi had been known since 1994. This much more complete skeleton presents a great deal of important information. A crucial behavior advancement with Ardi appears to be an omnivorous diet. This is suggested by its tooth enamel, which was only moderately thick. The thicker the enamel, the more equipped an organism is to eat plants, as they require a good deal of crushing and grinding to begin this first part of the digestive process. Meats are much softer foods. Other skeletal characteristics indicate that both tree-climbing and bipedalism were practiced. Given that these specimens have been found alongside faunal remains that indicate a wooded environment, it seems reasonable that Ardi was not a dweller of open savannah. Rather, they lived near enough to forests where their arboreal adaptations would be beneficial. This brings up an old debate in paleoanthropology – the woodland versus savannah hypothesis for early humans. That is, what is called the “Savannah Hypothesis” posits that human bipedalism evolved as a direct result of our ancestors transitioning from forests environments to savannah environments, where an arboreal lifestyle was no longer advantageous. Ardi would seem to suggest otherwise.
A Word About Bipedalism
There is no doubt that modern humans are primarily bipedal and that our Australopithecine ancestors were not. In between, there is a clear progression from walking on fours to walking on two legs. But, why?
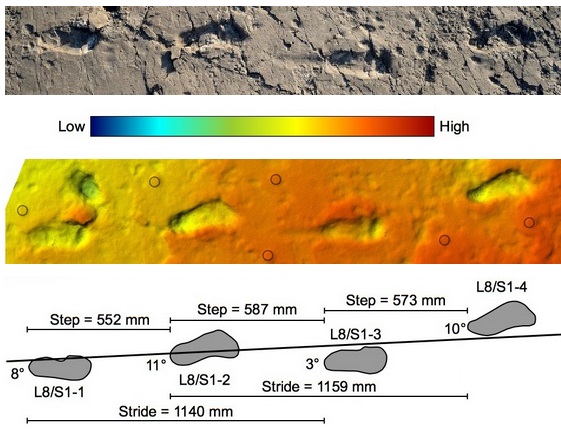
The advantages of bipedalism are very important. Bipedalism is a critical development that allows for the free usage of the hands for things other than walking or even climbing. Suddenly, carrying young is easier. Making tools becomes more possible. It also improves our long distance locomotion, making it easier to traverse much longer distances. Closely related to the ability to travel long distances is the ability to cool off, which bipedalism makes much easier, the body not being so hunched over on itself. Finally, with the climate changing so drastically in Africa, from tropical rainforests to savannah, being bipedal becomes a survival strategy. It is now possible to see over the grasses.
There are also disadvantages to walking on two legs. The curvature of the lower spine that makes balancing on two legs possible also leads to much more lower back pain. The move toward bipedalism led to the toe positions of the back feet being changed so that the big toe rests up against the next two. This makes “pushing off” while walking much more effective, but makes climbing trees like our ancestors very clumsy. Finally – and perhaps most importantly because of its role in reproduction – bipedalism works with a narrow pelvis. This narrow pelvis makes walking upright very efficient, but it makes childbirth much more difficult, given that the size of the birth canal is part of what is sacrificed for the ability to walk upright. In fact, in most modern human births, the head is too large for the birth canal. This is accommodated in female humans through the release of a hormone, relaxin, that softens the ligaments in the pelvis, allowing the various bones to create the space necessary for the birth to occur. This is augmented by the malleability of the baby’s skull, the bony plates not having yet fused. When all things don’t work together as they have evolved to do, birth complications arise. Modern medicine has found ways around such situations, with procedures such as Caesarean Sections, among others.
Here is an excellent video discussion of the wider variety of anatomical changes associated with hominin bipedalism.
A word about bipedalism for distance runners
Hominid bipedalism brought with it an ability to run. Running was not really as effective for our primate ancestors. It was more effective to use their arboreal, prehensile skills to climb trees. Why? Well, if you are escaping from predators or even chasing prey in a forest environment, these skills are quite useful.
But, what if the climate is changing and the environment along with it, so that tropical rainforests give way to savannah? How do you adapt and retain the ability hunt and escape – or just to travel long distances in search of food? The ability to run becomes important. And, not just the ability to run, but to run for long periods of time. Starting about 2Ma, the physiological ability to run, or really trot, become possible due to improvements in bipedal skeletal structure. These include the development of longer legs and shorter toes. Also, larger gluteal muscles, a smaller waist, and more neck flexibility played a part. The hominid foot had already also developed a larger heel bone (calcaneus) and longer Achilles tendons were in place. The ability to run also required changes in our metabolic abilities. These changes directly affected our brain, because both running and brain activity use a good deal of glucose. Changes in brain development that occur along with running include an enlarged anterior hippocampus, a change that likely led to improvements in memory (Schulkin, 2016). There are many more examples of physiological developments related to bipedalism from which we can draw in the literature. But, it is definitely worth pointing out here that while bipedalism itself brought important advantages, so did the endurance capabilities coupled with its evolutionary development for hominids.
Interestingly, sometimes evolutionary priorities conflict. In the case of the evolution of hominids, the ability to walk upright prevailed. This, despite the disadvantages that developed for childbirth, demonstrates that bipedalism must have been the more important survival strategy at the time.
Australopithecus anamensis, A. afarenesis, A. Africanus, A. garhi, and A. Sediba – 1.5-4Ma
These five species of australopithecines are all extinct, but provide important glimpses into the ancestral development of human features.
Australopithecus anamensis – 4.2-3.8Ma
Discovered near Lake Turkana, Kenya and also in Middle Awash, Ethiopia, A. anamensis shared traits we see in both apes and humans. Fossil evidence indicates bipedalism, particularly due to the human-like positioning of the ankle joint. But, long forearms also suggest that they climbed trees. Like Ardi, this specimen seems to argue for a still close connection to an arboreal lifestyle with some amount of bipedalism having developed. Its cranium also displays important features that connect it to humans, such as foreword projecting zygomaticus arch, or cheekbone. A. anamensis still retains important ape skull features, however, such as a narrow braincase and protruding face.

“Lucy” – Australopithecus afarensis – 3.85-2.95 Ma
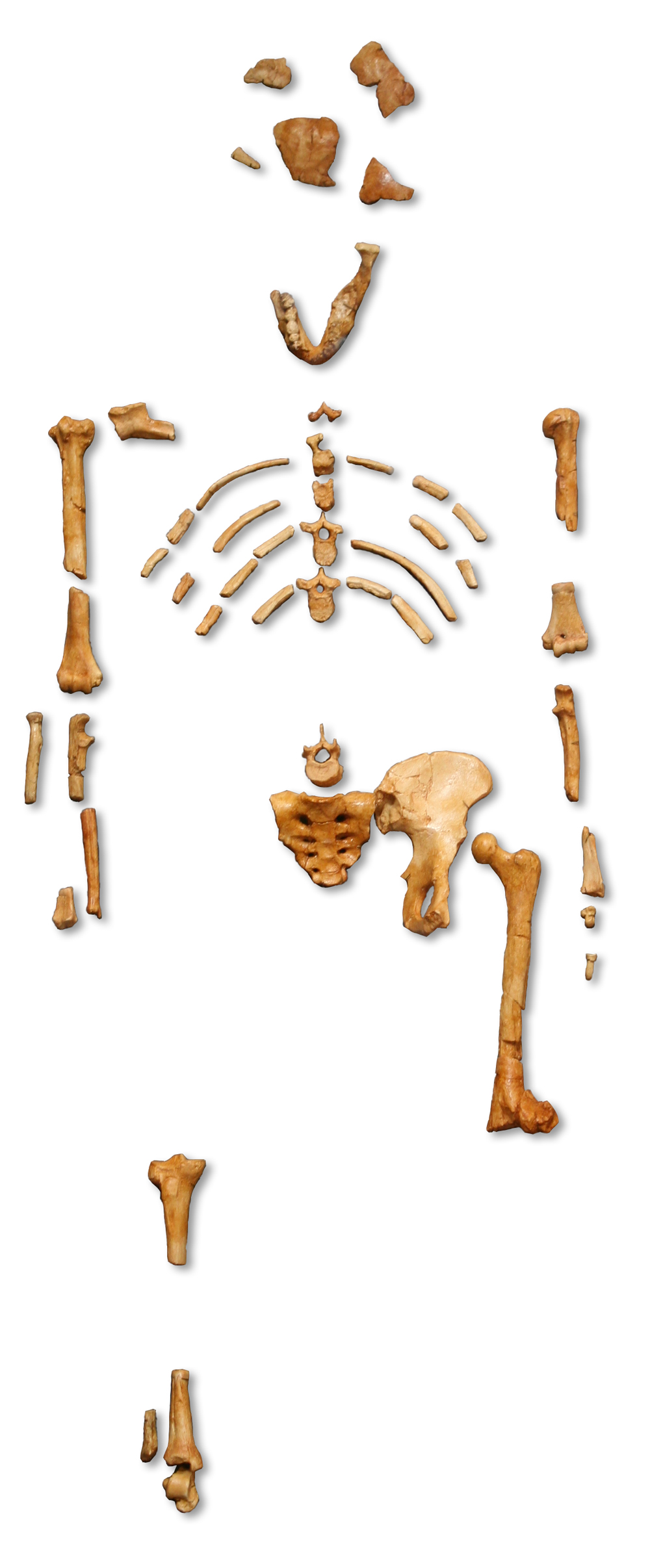
Image: 120, CC BY-SA 3.0, via Wikimedia Commons
Australopithecus afarensis is represented most famously by what is one of the most complete skeletons of an early hominid, nicknamed “Lucy”. There are over 300 individuals that comprise the fossil evidence of this species, so it is quite well studied. In many ways, this species represents a major signpost in human evolution, a place where we have enough information to confidently look both forward and backward when it comes to shared traits in our own species.
There is a fairly large degree of sexual dimorphism in A. afarensis, most notably in the difference in average height between adults males (151 cm) and females (105 cm). We know from skeletal evidence that, like chimpanzees today, they grew up into adulthood much faster than modern humans. Culturally, this would have left them with much less time for parental guidance and socialization during this brief childhood. Ian Tattersall (2012) has even suggested that they spent some of their times roaming savannah areas in herds like baboons are sometimes seen doing today, as opposed to living in small tribes like chimpanzees.
The species still retained many ape-like features, such as a flat nose and projecting lower jaw and a smaller braincase. It also retained the long, thin arms and long fingers that are well-adapted for tree climbing. Human characteristics come with smaller teeth and a body that stood fully on two legs and walked upright. It was an overall well-adapted species that a long-lived species, existing for nearly one million years.

Image: By Shalom – Own work, CC BY-SA 3.0
Note some of the skull features of A. afarensis. There are still ape-like protruding mouth and small brain case, but the occipital ridge above the eyes is much less pronounced and the zygomaticus arch (cheekbone) is more forward positioned, as is the foramen magnum on the bottom of the skull. Dentition not shown. (Model: Smithsonian Institution, Creative Commons License)
Did I Get It? Bipedalism
Australopithecus africanus – 3.3 to 2.1 Ma
A. africanus is very similar to its immediate ancestor A. afarensis. Distinguishing features include a more rounded cranium with a larger brain and smaller teeth. Based on their teeth, it is thought that they were vegetarian, unlike some of their ancestors. For a long time, they were viewed as predators. This was not because any discovery of stone tools, but because some specimens had been discovered with broken animal ones nearby. Since then, it is now thought that these represent animal predation on A. africanus, rather than the other way around.
Still, these individuals were fully bipedal and yet were still well adapted for arboreal life. Specimens have been found primarily in South Africa, which may indicate this species was a southern cousin to A. afarensis.
Above: Cranial specimen of A. africanus. There are many still-developing human features as described in above examples, but the larger braincase here is probably the most important development.
Above: Cranial specimen of A. africanus. This specimen allows some examination of dentition. Note the lack of a protruding, large, canine tooth, something that would be indicative of more ape-like dentition. The flat molars presented here are indicative of a vegetarian diet. (Model: Smithsonian Institution, Creative Commons License)
Australopithecus garhi and A. sediba
Of all of the Australopithecine specimens, these two latest-lived species are poorly known, as well as short-lived. A. garhi is only known from a few fragments. But, because it dates to a very poorly-documented time period in human evolution, 3-2 Ma, it is very important. The few fragments that do exist indicate a significant increase in bipedal stride length, and thus a more efficient gait. Another important piece of evidence are the stone tools that have been found in association with A. garhi. One of the first documented examples of tool usage among early humans, the fragments are solid evidence of hunting as a form of food procurement.
A. sediba is known from a very well-preserved and nearly complete skeleton found Malapa Cave, South Africa. This species lived at the same time that the genus Homo first appears in the fossil record. At the tail end of the Australopithecines, A. sediba lived alongside some of our earliest direct ancestors.
This detailed skeleton reveals some important developments. Changes in the pelvis indicate more upright walking, various tooth traits and a broad chest are more human-like, features of the femur and legs overall indicate that it spent a much greater time walking and doing so in a very similar fashion to modern humans, much more so than its other Australopithecine ancestors. Derived features of its skull are also more human, such as small premolars and molars and facial features that are less pronounced and more human.
A. sebida skull. Note the more human features, such as the larger braincase, less forward protrusion in the lower jaw, and a more compact dentition. 3D Model: Creative Commons, William Mitchell
The Genus Homo – 2.4 Ma
The genus Homo appears to have split from Australopithecines somewhere between Australopithecus africanus and Australopithecus garhi. The earliest direct ancestor of the genus is represented by Homo habilis, a species known best from specimens found in parts of eastern and southern Africa between 2.4 and 1.4 Ma.
It is very important to note that the early specimens of the genus Homo are all found in eastern Africa. Very rapidly after the evolution of Homo erectus, sometime around perhaps 1.5 Ma, human ancestors are found in other parts of the world than just Africa. At that point, the human diaspora had begun.
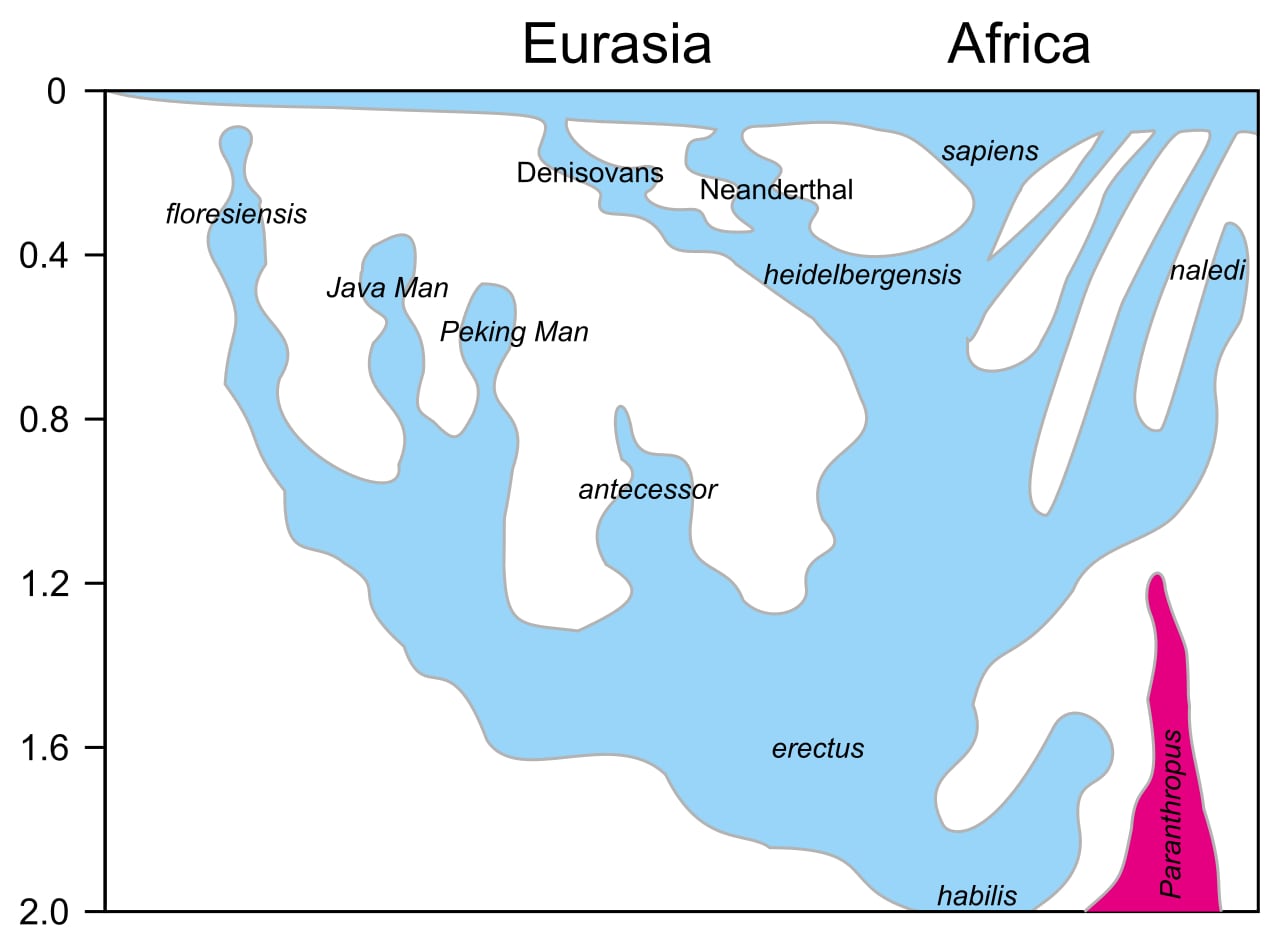
Image: Conquistador, User:Dbachmann – updated version of File:Homo-Stammbaum, Version Stringer-en.svg, CC BY-SA 4.0
Homo habilis – 2.4-1.4 Ma
Our earliest Homo ancestor, Homo habilis, had a varied diet that appears to have been omnivorous. Originally discovered by the Leakey family — Jonathan Leakey specifically — the early specimens were found in Olduvai Gorge, Tanzania. It is the first of the genus Homo to be associated with the use of stone tools, due to the massive amounts of flakes and tools found in context with the skeletal remains. Later, cut and smashed animal bone would also be found in context with H. habilis, suggesting that meat was important to the diet on some level.
A Word About Meat and Brains
Since the 1950s, paleoanthropologists associated increased cranial capacity to the increasing meat content in the diet of human ancestors. Evidence supporting this hypothesis include that Homo habilis was a known toolmaker and those tools were used in some part for meat procurement. Additionally, up until H. habilis, hominid cranial capacity is increasing. Discoveries such as these in ancient Ethiopia, around what was then a rather verdant lake, have been interpreted as the influx of protein that led to major evolutionary and cultural advances in the human lineage. Yes, meat-eating made us smarter. Humans are the only primate that eats other large animals other than those of our own species (and cannibalism is well-known, too). Butchery marks on animal bones, stone tools, and the genus Homo all come around the same time in the fossil record.
However, new research suggests that, rather than the meat, it might have been bone marrow and associated nutritious fats that helped early human brain development and tool development (Thompson et al., 2019). Early humans may have eaten meat, but eating meat out in the open was a dangerous business for our ancestors, as other predators and scavengers lurked nearby, and the meat itself was difficult to move. And, with so many very large mammals, it is more likely that humans were some of those scavengers, opportunistically awaiting while large predators stripped hunted carcasses of meat, leaving behind bones. Bones could be then be removed and easily carried to safe locations for consumption. Further, meat quickly rots, while bone marrow stays fresh longer. The geologic epoch of this transition, the Pliocene, was also a time of major environmental change, as the climate was cooling intensely and was becoming more variable. Large swaths of what was rainforest were becoming grasslands and savannah. After 2 Ma, human ancestors were likely consuming meat in addition to bone, but it is very likely that bone scavenging and processing with stone tools came first. The conclusion is that we ate marrow before we ate meat.
Here is a nice overview of some of the ongoing discussions of the ancient dietary changes in our hominid ancestry at this point.
Homo habilis skull, where many early human, yet still very ape-like features can be seen. Human features include a larger cranium, lack of a protruding canine tooth, more compact dentition due to a less protruding mouth, and a downward/forward foramen magnum. There is still a very pronounced occipital ridge above the eyes. 3D model by BuluLulu on Sketchfab.
Next, examine this chopper tool from Olduvai Gorge, Tanzania. At 1.8 Ma, this tool could have been used by Homo habilis to access bone marrow. Tools of this kind were often very stylized, an indication of common culture, as uniformity in stone tool manufacture indicates not only a transfer of cultural skills and knowledge, but also, potentially, specialization of skills to some degree within these early ancestral human populations. Flakes were removed form a larger core to create a sharp edge. This edge could be sharpened further until it was no longer of use.
Stone scraper, and early stone tool. (Model: Smithsonian Institution, Creative Commons License)
An excellent introduction to stone tool technologies and their development can be enjoyed in the video below.
tools
We know that human ancestors are not the only animals to use tools. One brief example outside of our human family are chimpanzees. Jane Goodall famously documented chimpanzee tool usage while living among chimpanzee communities. Behaviors she observed included picking up leafy twigs, stripping the stems of the leaves, and using these stems to ‘fish’ for termites and ants. Since then, chimpanzees have been seen using tools to crack nuts as well as sharpening sticks to hunt for other mammals. In the last thirty years, gorillas and bonobos have also been observed using tools of various kinds.
Primates are not the only tool-users either. Birds, whales, sea otters, and even various predators have been observed using tools of various kinds. Given all of this, tool usage is certainly not indicative of human behavior, but something hominids likely inherited as an ancestral adaptation, but seems to be a behavior that has developed independently, evolving in multiple directions.
Did I Get It? Intelligence
Homo erectus and Homo floresiensis– 1.89 Ma to 50 ka
Homo erectus has been well known since its first fossil discovery in Indonesia in 1891. This discovery actually predates all discoveries of human ancestors in Africa. The discoverer, Eugene Dubois, originally named the species Pithecanthropus erectus, or “erect ape-man,” but it was popularly referred to as “Java Man.” Many discoveries of this species would follow, both in China (where it was called “Peking Man”) and in east Africa.
Homo erectus was quite humanlike in its proportions. By this point, it had lost the long arms and prehensile ability of its arboreal ancestors. These traits had given way to a modern human gait and stature. H erectus was on average rather tall, ranging from 145-185 cm (4.7-6 ft), with an associated weight range of 40-68 kg (88-150 lb). It is very likely that not only did they spend most of their time walking, but running was likely important to their locomotion, possibly for long distances. Young appear to have grown quickly, like other apes, but the species has shown evidence that it may have cared for elders and weaker individuals. Homo erectus is also associated with a major stone tool development, the hand axe.
Homo erectus skull. Note the much more human-like characteristics, despite the still pronounced occipital ridge. The face protrusion is much less pronounced than in its ancestors and the cranial capacity is much greater. (Model: Smithsonian Institution, Creative Commons License)
Above: A hand axe dating back to 1.1Ma from Isampur, India, that is associated with use by Homo erectus. Stone tools like these are much more advanced in design and in the skill necessary to create them, indicating the development of some specialization in this craft. (Model: Smithsonian Institution, Creative Commons License)
Flint-knapping, or the hand-shaping of rock tools, was not easy. It required highly specialized and learned skills, particularly as the work progressed from simple scrapers to well-hewn spearpoints. It is very difficult. The video below provides an excellent introduction to the ancient set of skills:
Video of Tyson Hughes demonstrating flint-knapping (stone tool manufacture) for Crow Canyon Archaeological Center, Cortez, Colorado.
Homo erectus is certainly the longest-lived species of Homo. It lived for longer than anatomically modern humans have yet managed to exist. Their tall frame and large brain required a very large amount of energy to function, making meat-eating for easily digested protein a key ingredient in their diet. Their stone tool development certainly points toward the processing of meat as being an important part of the diet. One other development that is important to point out at this point is the use of fire. Up until Homo erectus, there is no evidence in human ancestry of the purposeful starting and use of fires. Now, the remains of hearths are found in context. While these were almost certainly used for cooking, they were also apparently used for social interaction, warmth, and to keep predators at bay.
Here is a great video showcasing some of our developmental history with fire.
An extinct descendant of Homo erectus is Homo floresiensis. Nicknamed “the hobbit”, it has only been found very recently (2003) on the Indonesian island of Flores. H. floresiensis is also associated with stone tool use, but it appears to have gone extinct as late as 50 ka. Known individuals (and there are only a few) stood only 106 cm (3.5 ft) tall, had tiny brains, large teeth, and an array of unusual facial features. These include flat foreheads and less pronounced chins. Their feet were very large for their legs as well. Despite all of this, there is evidence that they subsisted in part on small elephants, large rodents, used fire, and made stone tools. Large monitor lizards (like Komodo dragons) may have preyed on them. One possibility for the distinct stature and overall form of H. floresiensis is island dwarfism.
Homo heidelbergensis-1.3 Ma-200 ka
Eventually, human ancestors would spread beyond Africa, China, and Indonesia and into Europe. The first specimen of Homo heidelbergensis was discovered Near Heidelberg, Germany, in 1908. This is the first species of ancestral human to live in cooler climates. Living in Europe in the midst of the Pleistocene epoch’s “Ice Ages” would not have been simple, so we can hypothesize a need for body coverings, skill with fire, and other cold-weather adaptations, including the hunting of large animals. Indeed, we have much of this. There are fire-altered tools and wood at a the site of Geshre Benot Ya-aqov in Israel, for example. H. hiedelbergensis also likely built shelters. Evidence of this comes from the site of Terra Amata, France.
Evidence for their hunting of big game comes from analysis of the bones of those animals where they have been found in context with H. heidelbergensis remains. These include butchery marks on deer, horses, elephants, hippos, and rhinos. An example of a site with such features as an early thrusting spear exists near Schöningen, Germany.
Finally, there is evidence of human ritual among H. heidelbergensis near Atapuerca, northern Spain. An apparently purposeful mass burial of 30 individuals dates to 400ka. Was this ritual? Certainly, there was some kind of purpose in such a burial. This pit, nicknamed Sima de los Huesos, or “Pit of Bones” also contained evidence of stone tool use, as a hand axe was also found in context.
A skull from H. heidelbergensis. Note how human this skull appears. The braincase is much larger, and the face much flatter. The cheekbones are less pronounced. There is, still, a rather elevated occipital ridge, which is a more ape-like feature shared with all ancestral humans up to this point. (Model: Smithsonian Institution, Creative Commons License)
Homo naledi – 335-236 ka
Remains of Homo naledi has so far only been found in South Africa. H. naledi are cousins of humans who never left Africa. At the site where this species has been found, there are 15 individuals that have been recovered. It is one of the largest single fossil hominid caches known. So, there is a fair amount of physiological information within that small population, including both adults and juveniles. Their remains have been dated to between 335,000 and 236,000 years of age.
Despite clear similarities with other species of the genus Homo, a number of features from Australopithecus are also retained. This has made the placement of H. naledi on the hominid phylogenetic tree challenging.
Neanderthals
Until now, the only one species of Homo has been known to have migrated out of Africa, Homo erectus. We have little information on the fate of that species, but we do know that it died out around 117,000 years ago, well before modern humans, Homo sapiens, would encounter some of the same environments in Asia. Thus, there is currently no evidence that these two species ever interacted.

Image: Wikimedia, By Bacon Cph CC BY 2.5

Wikimedia, Pressebilder Neanderthal Museum, CC BY-SA 4.0
The situation with Homo sapiens neanderthalensis is quite different. Neanderthals are a subspecies of our own species, and there is ample evidence that they also originated in Africa. They left the continent well before our species, Homo sapiens sapiens would begin its own migration (350 ka). Once they left, they migrated throughout Asia and into Europe. Given this timeline, it is possible that they interacted with Homo erectus individuals living in Asia at the time, though at present, there is no decisive evidence confirming this. Recent information suggests that ancient close relatives of Neanderthals, called the “Denisovans,” lived alongside and interbred with our species for millennia. Known mainly by their unique DNA variants found in modern humans, they outlasted their Neanderthal relatives by at least 10,000 years, only going extinct as recently as 15,000 years ago. The last documented evidence of a co-existence between Denisovans and modern humans, placed via DNA evidence at 30 ka, comes from New Guinea (Jacobs et al., 2019).
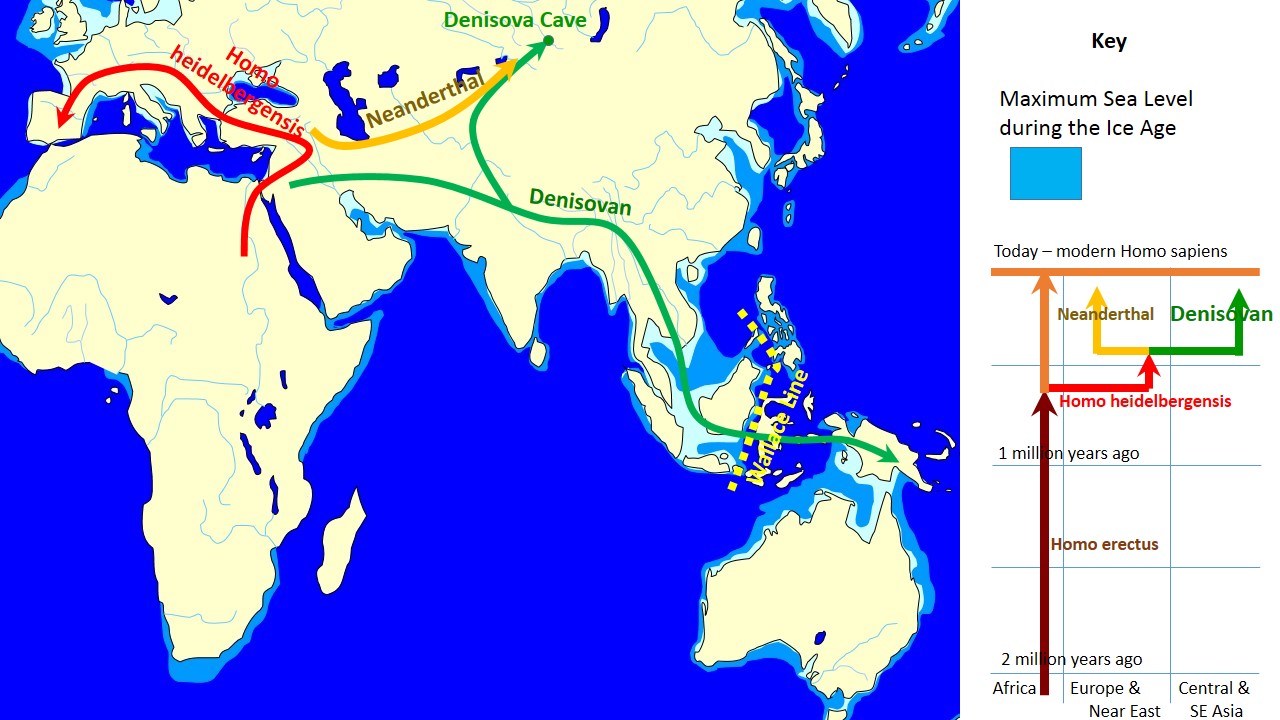
Image: Wikipedia, John D. Croft, CC BY-SA 3.0.
Modern humans and Neanderthals did briefly interact in Europe before Neanderthals would also go extinct there. It is not known why Neanderthals went extinct, though a changing climate and the various natures of their interactions between modern humans have been suggested as hypotheses. Some examples of the latter include interbreeding with humans (being assimilated into the Homo sapiens sapiens population) and birth problems, disadvantages with humans in the same ecosystems, parasites, pathogens, and violence. One of the last known occupations of Neanderthals in Europe or Asia is found in a cave on the coast of Gibraltar, Spain. Remains there are dated to 42 ka.
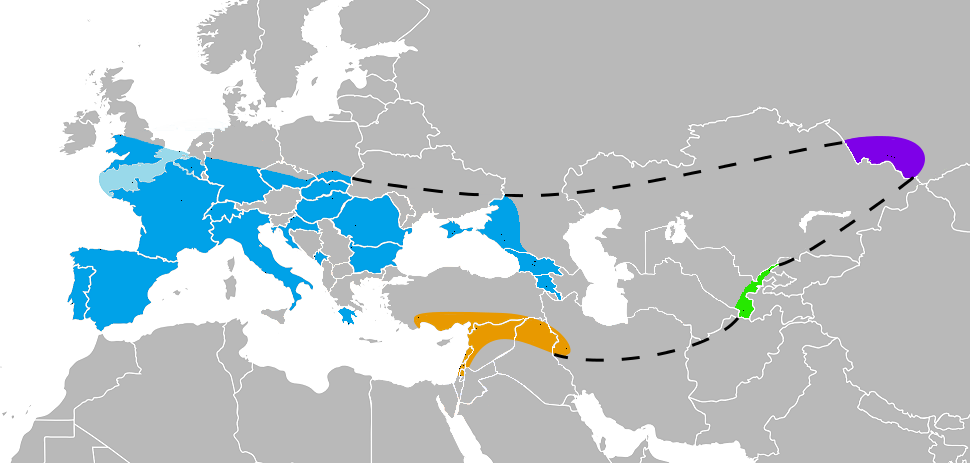
Image: Wikimedia, CC BY-SA 3.0
Upper portion of the skull of a Homo sapiens neanderthalensis. All of the features consistent with modern humans are present, save for a thicker occipital ridge. (Model: Smithsonian Institution, Creative Commons License)
Neanderthal lower mandible showing dentition that is very consistent with modern humans and a mixed omnivorous diet. Preserved mandibles are more rare in fossil hominids. This mandible is included here as an example of mandible dentition in Neanderthals, as they have lost any pronounced lower jaw canine as well as the associated diastema. (Model: Smithsonian Institution, Creative Commons License)
An entire chapter could be devoted to what we know about Neanderthals. In terms of their culture, we know that they had a diverse toolkit, built shelters, harnessed fire, used clothing, hunted large game (they were apex predators), ate plants, and also participated in ritualistic behaviors. This last point is most significant. No other prior hominid shows the degree of ritualism known among Neanderthals, other than modern humans. An excellent example of this is the inclusion of what appear to be offerings such as flowers for the dead who have been buried.
In lieu of a long exposé on Neanderthals, the video below highlights some additional information that is helpful in helping us understand our own humanity, through the lens of their fossils.
Also unique to Neanderthals is the existence of actual direct DNA samples from several individuals from a wide variety of locations, including Siberia, Croatia, Spain, and Germany. By 2009, the entire known sequence of Neanderthal DNA had been sequenced by the Max Planck Institute for Evolutionary Anthropology, as a part of the Neanderthal Genome Project.
Modern Humans
This case study has been documenting your story and my story. It is a story of our hominid ancestors in deep time. The full story of modern humans is varied, complex, and beyond the scope of this case study. However, there are a few important things to point out:
Like all other hominid species, we emerged from Africa. This migration began between 100 ka and 60 ka, both along the Sinai Peninsula of Egypt and across what is now the Bab-el-Mandeb Strait between Djibouti and Yemen. At the time of migration, the last glacial period was approaching, ice sheets were advancing to the north, and sea levels were much lower. Parts of Africa that were covered with tropical rainforest were giving way to Savannah and even desert, as a part of what is sometimes referred to as a megadrought (Gibbons, 2007). Essentially, our ancestors, Homo sapiens sapiens, were driven to greener pastures to the east, north, and further south deeper into Africa.
While there is ample fossil evidence for humans having left Africa at this time and then having migrated throughout Asia, Europe, and into North America (across Beringia as early as 16.5 ka), it is difficult to trace this migration in detail. However, multiple studies have attempted to fill in these gaps using DNA by tracing mitochondrial DNA (on the maternal side) and via the Y-Chromosome (on the paternal side). These studies have traced all modern humans back to a “Mitochondrial Eve” who lived in east Africa about 150 ka. Based on this DNA, maternal haplogroups (individuals who share a common ancestor) can be traced across the world, including into North America. All females, even members of groups who never left Africa, share this mitochondrial signature.
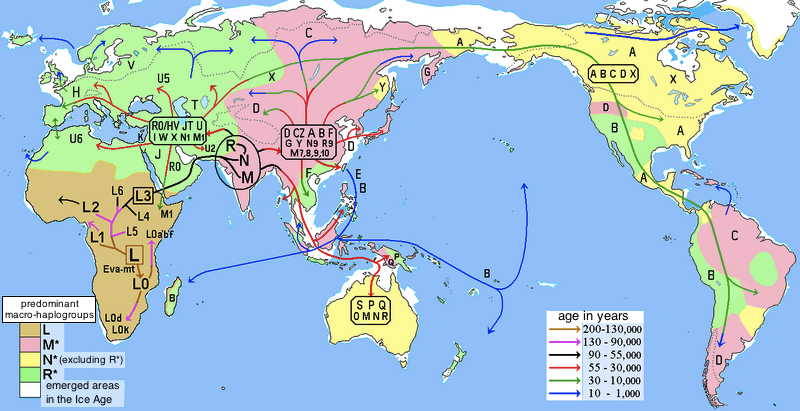
Image: Wikimedia, Maulucioni, CCBY-SA 3.0
Likewise, paternal haplogroups can be traced (using the Y-chromosome) across the world. In this case, however, all men who share this version of the Y-chromosome are descended from a single man who appears to have migrated out of Africa about 60,000 years ago, likely taking the southern route across Yemen to do so. It’s important to note that some human populations, such as members of the !Kung people of Namibia, Angola, and Botswana, do not share this Y-chromosome. This is because they are descended from other paternal line that split away prior to 60,000 years to venture deeper into western Africa.
The map below is an ArcGIS Online layer that illustrates the migration patterns of various maternal (mitochondrial) and paternal (Y-Chromosome) haplogroups among Homo sapiens sapiens as they migrated out of Africa. Clicking on the boxes toggles on and off the various haplogroups and you can see where members of that group migrated around the world:
Instructions for how to use this map can be accessed here.
Haplogroup.info, used under CCBY 3.0.
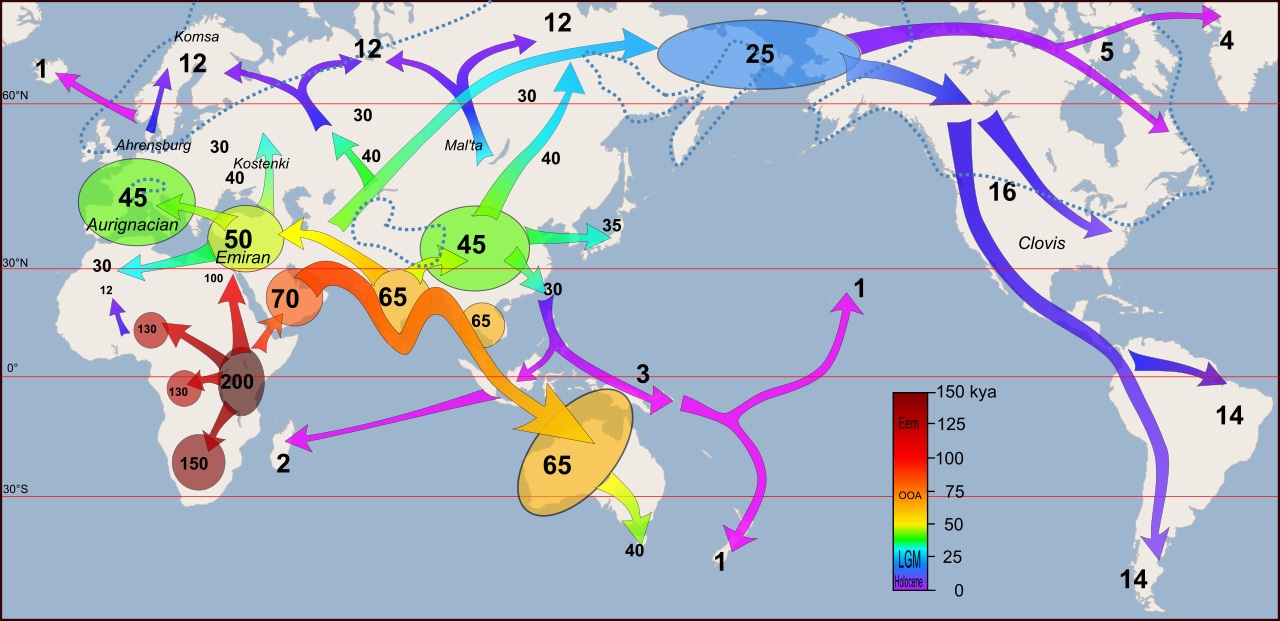
Image: Wikipedia, DBachman, CC BY-SA 4.0.
We can also trace this migration, particularly the latter portions of it, using an important evolutionary and cultural development: language. Like evolutionary change among isolated pockets of species, language development is rapid, very culturally specific, evolutionary in nature, and is thus a very useful tool for anthropological and archaeologic studies of modern human migration and culture. There are a wide variety of language groups across the world, all of which can be traced by linguists to a handful of primary language components that, combined, reveal aspects of what once may have been a “mother tongue” for that primary language family.
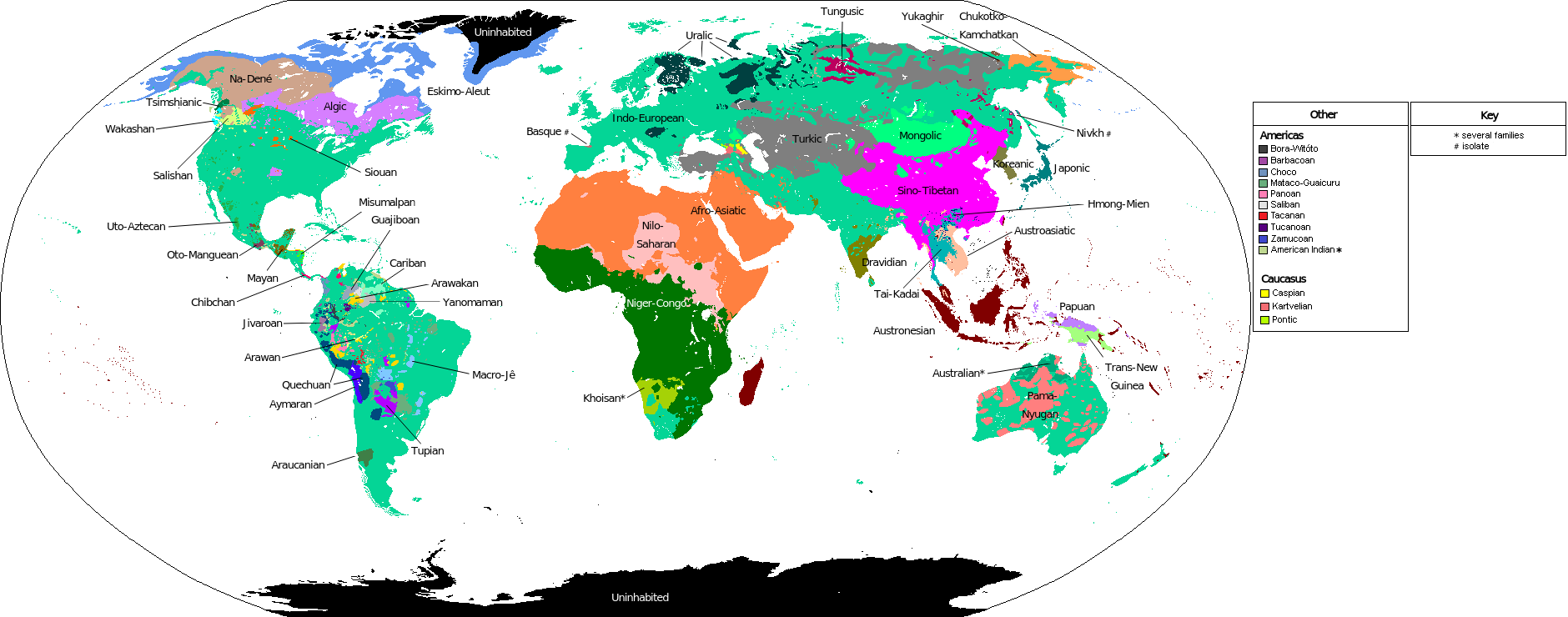
Image: Wikipedia, Alumnum, CC BY-SA 4.0
Image: Wikipedia, Joshua Jonathan, CC BY-SA 4.0
You can view a video animation of the spread of Indo-European and Uralic language group speakers below:
Did I Get It? Haplogroups and Migration
Homo sapiens, Climate, and Natural Resources
What caused all of this migration? Homo sapiens sapiens, like all other animals, seeks to eat, procreate, and survive. Unlike many other animals, our species migrated in ways that go beyond some kind of seasonal round. We have been able to adapt through our use of tools, our diet, and in the pursuit of resources. Today, our species lives far differently (in most cases) than our ancestors did thousands of years ago and, for most of us, we live very differently than our recent ancestors just a century or even a few decades ago. What has driven these changes?
A significant part of that answer lies in climate change. Before we actively played a role in altering our climate, as we do today, our climate dictated where we called home to a large extent. Climate change contributed as a trigger for migration in many cases. Despite this, our ancestors did brave much colder climates and the adaptability challenges those climates brought with them. Technological advances have occurred irrespective of just those driven by climate, however. While certainly adapting to colder areas required innovation of our toolkits, we innovated these kits in warmer climates also. The Stone Age would give way to Neolithic technologies (agriculture, animal husbandry). To these new Neolithic ways we would eventually add our harnessing of metals such as copper, bronze, iron, and eventually silicon (today). All of this innovation requires further resource extraction. Fundamentally, this resource extraction is similar to that practiced by our microbial ancestors over a billion years ago. As with simpler organisms, all human uses of our environment have been toward the goal of providing our species with food, shelter, and a chance to reproduce.
We’re animals, after all.
Did I Get It? Humans and Climate
Further Reading
Gibbons, Ann (2007). Megadrought put the squeeze on our ancestries. Science News.
Jacobs, Guy S., Hudjashov, Georgi, Saag, Lauri, Kusuma, Pradiptajati, Darusallam, Chelzie C., Lawson, Daniel J., Mondal, Mayukh, Pagani, Luca, Ricaut, François-Xavier, Stoneking, Mark, Metspalu, Mait, Sudoyo, Herawati, Lansing, J. Stephen, and Murray P. Cox (2019). Multiple deeply divergent Denisovan ancestries in Papuans. Cell 177(4).
Patterson, Nick, Richter, Daniel J., Gnerre, Sante, Lander, Eric S., and David Reich (2006). Genetic evidence for complex speciation of humans and chimpanzees. Nature 441(7097).
Schulkin, Jay (2016). Evolutionary basis of human running and its implications on neural function. Frontiers in Neuroscience 10:59.
Smithsonian National Museum of Natural History Human Evolution Timeline, Accessed 2/4/2021.
Smithsonian National Museum of Natural History Human Origins Initiative, Accessed 2/4/2021.
Tattersall, Ian (2013). Masters of the Planet: The Search for Our Human Origins. St. Martin’s Griffin.
Chapter Contents
- 1 Humans as a species
- 2 Paleoanthropology
- 3 Highlights of our family tree: Australopithecina and Homo
- 4 Further Reading