KEY CONCEPTS
- Dinosaurs have many lines of circumstantial evidence that implies social groups, like mass graves and footprints
- There is still no direct evidence of pack hunting, but there is direct evidence of other hunting strategies
- Dinosaurs have strong evidence that at least some species had intensive care of their young, including nests and parental help
- Many dinosaurs might not be separate species or valid due to incomplete remains, sexual dimorphism, and changes to animals as they age
- Famous examples include Brontosaurus/Apatosaurus, Triceratops/Torosaurus, and Pachycephalosaurus/Stygimoloch/Dracorex
- Though not every fossil shows impressions of skin or feathers, enough have been found to show how they developed in theropods
- Recent studies have shown a larger number of species had feathers, though large animals like Tyrannosaurus rex probably were still featherless
- Dinosaurs have significant evidence that they were not ectothermic (cold blooded), and may have been endothermic (warm blooded)
- Feathers and their size may have helped dinosaur thermoregulation
- At the very least, dinosaurs were agile and active animals who could move much more like mammals and birds than lumbering lizards as they were presented prior to the dinosaur renaissance
Dinobook or Saurogram? – Dinosaur Social Behavior
There is a lot of debate over the social-networking skills of dinosaurs. On one hand, the living descendants of dinosaurs (birds) are highly social. On the other hand, social interactions are inherently difficult to preserve in the fossil record, so direct evidence is hard to come by. The two main ways that social behaviors can be inferred is through trackways and mass mortality sites.

Dinosaur track sites are known in many places around the world. Lots of useful information can be gained from them, including speed and gait. Some of them have enough tracks to represent multiple individuals and even multiple species. When this happens, paleontologists can infer social behaviors that would never be preserved otherwise. Each dinosaur group has a characteristic style of tracks, though in many cases, the exact dinosaur that laid them down is hard to distinguish. Trace fossils like tracks are also given their own nomenclature that follows the same rules as naming a biologic organism, complete with geniuses and species.

A good example of a dinosaur track site which has a reasonable interpretation at behavior is the Ardley Quarry in the UK. It not only shows a large group of animals walking the same direction (including possible prey and predators), but even different ages all together, similar to what is seen in large mammal herds today. This does not prove that dinosaur herds and modern mammal herds had the same structure, but it is a good indication. It has even been used to make evolutionary inferences about the extent of radiation of different dinosaur groups.
Another famous example is Dinosaur Stampede National Monument in Australia. There is still debate as to what exactly happened at this site, where a lone large individual was walking through the mud with many different smaller individuals. Regardless of the exact circumstances, the large variation in the smaller tracks give an indication that the group was a herd with different-aged animals.
Mass mortality sites also help get at the behavior of dinosaurs, with the general thought that if they lived together, they died together. The first of these was discovered in Belgium in 1878 when nearly 40 articulated Iguanodons were found. In 1974, at least 14 Pachyrhinosaurus individuals were found together in an ancient Alberta riverbed, most likely washed up together when trying to cross a flooded river. There are countless others, but just to highlight some important ones: In 2008, a find in Inner Mongolia, China was announced that had more than 20 Sinornithomimus animals that were all immature sub-adults, between one and seven years old. It is proposed that they were a social group, perhaps left to their own devices while adults cared for the infants.

Deinonychus, the dinosaur that the raptors from the Jurassic Park movies were based on, has a lot of interesting evidence of social behavior. North American fossil sites have found a lot of individual animals of varied ages in concentrated sites. Also, they have been found in close proximity to a possible prey animal, called Tenontosaurus. Deinonychus was about 1/20th the size at best, so how did they bring them down? Many have speculated that this is an indication of pack hunting. Certainly, their proximity in the fossil record could have been related to scavenging, but pack hunting is an intriguing interpretation.

The most unambiguous dinosaur fossil depicting behavior is the famous fighting dinosaurs. Found in the Gobi in 1971, it is a fossil that has a Protoceratops and a Velociraptor (the smaller relative of Deinonychus) forever locked in combat. The Velociraptor has its killing claw lodged into the side of the Protoceratops, while it fights back by grabbing the attacker’s arm in its beak. There is nothing yet found that comes close to depicting behavior like this fossil, which is why many consider it to be the greatest find ever.
Rearing behavior
Another aspect of social behavior is parental investment. Many reptiles are not known for being great mothers and fathers, with sea turtles famously laying their eggs on a beach and leaving. The closest relatives of dinosaurs (crocodiles and especially birds), however, are noteworthy parents in the animal kingdom. So where do dinosaurs fall on this spectrum?

The first noteworthy discovery on this front was nests. In the early 1920s, an expedition led by Roy Chapman Andrews in Mongolia found what is believed to be the first dinosaur nest and eggs. Near the nest was a skeleton, described and named by Henry Osborn as Oviraptor. Initially, the nest was thought to belong to a common dinosaur in the stratum Protoceratops. Oviraptor, which means “egg thief,” was assumed to be stealing the eggs and eating them with its toothless beak. Even Osborn at the time cautioned the use of such an interpretation in its name, but it stuck. In the 1990s, more discoveries, like the related animal Citipati found fossilized in a brooding stance over a nest helped clear the image of Oviraptor, though it is too late to change the name.

Maiasaura, the “good mother lizard,” with the femanine ‘saura’ used for “lizard,” was described by Jack Horner in the 1970s. Its discovery has done more to advance our understanding of parental behavior than any other dinosaur. Instead of finding just one dinosaur or one nest, an entire nesting ground was discovered, spaced closer together than the length of one adult, as some birds do today. The eggs were arranged in a spiral and placed on top of rotten vegetation to keep them warm (instead of using the parents’ warmth). The most important find was of hatchlings, which had underdeveloped legs which made walking unlikely, and yet, had worn teeth. It is estimated that the hatchlings might have spent a year in the nest, and although the mortality rate was high, it was the parents of these babies that fed them and protected them.
A Dinosaur by Any Other Name – Species controversies

When a paleontologist finds a new fossil, one of the first things they analyze is the dimensions of the fossil to compare it to existing species. If the organism is different enough, then a new description is written about it, defining the new species. Seems easy, right? There is a problem though. How does the paleontologist know how representative the organism was to its species? Imagine you are a future scientist studying human fossils. If you came upon the bones of a male basketball player, and then the bones of a female gymnast, would you think they were the same species? How about an elderly person versus a juvenile?

Of course, part of the training of any paleontologist is studying modern anatomy and populations of organisms so you have a basis for comparison for things like sexual dimorphism (changes between the genders of a species), gerontology (changes based on age), variability within a species, or even diseases that may alter the appearance of tissues. But when presented with an extinct species, certain extrapolations and assumptions are required. This has led to several controversies in paleontology, notably within dinosaurs. In fact, a very large number of dinosaurs, even some famous ones, may be nothing but a synonym of an existing species. In biologic nomenclature, the first name published gets precedent. This is why Ultrasaurus, Nanotyrannus, and Monoclonius are now considered invalid, and some scientists feel as many as one third of dinosaur species may join them in infamy. Some other famous examples are described below.
Brontosaurus
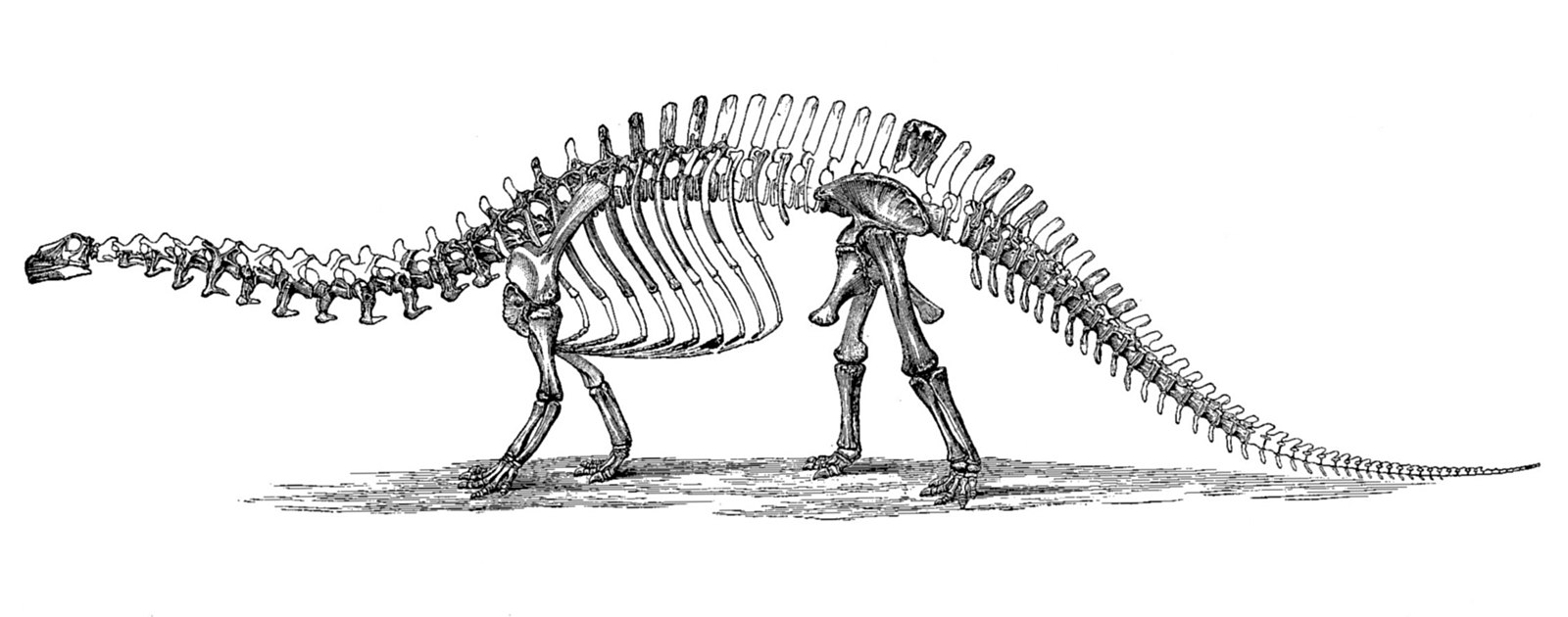
Perhaps the most famous dinosaur naming faux-pas comes from Brontosaurus. It is still one of the most famous dinosaurs ever, but it might not even exist! Toward the end of the 19th Century, scientists O.C. Marsh and Edward Cope were in a constant battle to describe and discover as many dinosaurs as possible, a rivalry known as the Bone Wars. This led to rushed, inaccurate, and sloppy science as they fought for prominence in paleontology, and Brontosaurus was one of the victims. In 1877, Marsh described a sauropod which he named Apatosaurus. Two years later, Marsh described a similar animal, but different enough in his mind to warrant a new genus. He named it Brontosaurus. One part of the skeleton was missing, however: the skull.

In 1903, Elmer Riggs, a paleontologist from Chicago’s Field Museum, said that Apatosaurus and Brontosaurus were similar enough to be the same genus, and since Apatosaurus was named first, the term Brontosaurus became invalid as a junior synonym. However, when the first mount was made for the American Museum of Natural History (the first sauropod mount ever displayed) in 1905, the name Brontosaurus was chosen for the skeleton. Since the skull was not known, a bulky skull similar to Camarasaurus was used. This mount may at least partially explain why the term Brontosaurus has persisted, even though throughout the 20th Century, almost all scientists agreed with Riggs that Apatosaurus was the proper name that should be used. In 1909, an Apatosaurus skull was found (which was more slender, similar to another sauropod, Diplodocus), but the knowledge of this skull remained limited until it was published in the 1970s.
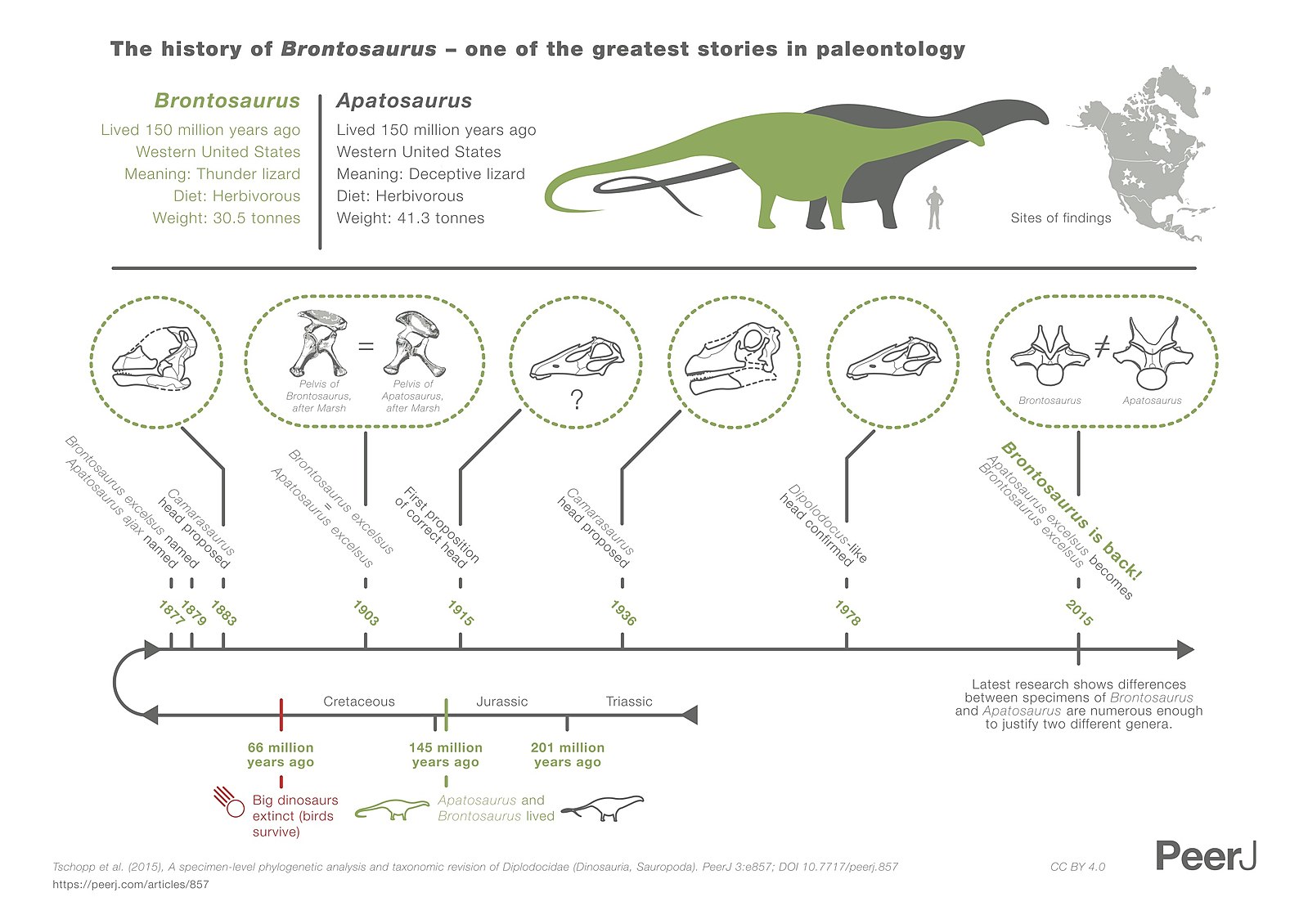
In 1998, Robert Bakker argued that different species of Apatosaurus should be split into multiple genuses. Then, in 2015, Emanuel Tschopp, Octavio Mateus, and Roger Benson released a study on Apatosaurus and related animals and their analysis also concluded that there is enough variation in the Apatosaurus’ to justify a new genus name. Rather than create a new name, they decided to resurrect the Brontosaurus name for this new genus. While not all paleontologists agree, it may have solved the problem of the popular, but potentially incorrect, Brontosaurus.
Toromorph

One of the more infamous controversies is called the toromorph hypothesis. Triceratops is a famous ceratopsian dinosaur from the late Cretaceous of western North America. Torosaurus is from the same general time, place, geologic formation, and habitat. It is noticeably larger than Triceratops, with a larger frill with large holes in the center of each half of the frill. Starting in 2009, John Scanella and Jack Horner proposed that Triceratops was just a juvenile of Torosaurus. Other related animals, like Nedoceratops and Ojoceratops have also been suggested as growth stages of Triceratops. Evidence to support this position includes several juvenile-like features of Triceratops, dramatic changes in Triceratops’ known young versus known ‘adults,’ and a lack of Torosaurus young.
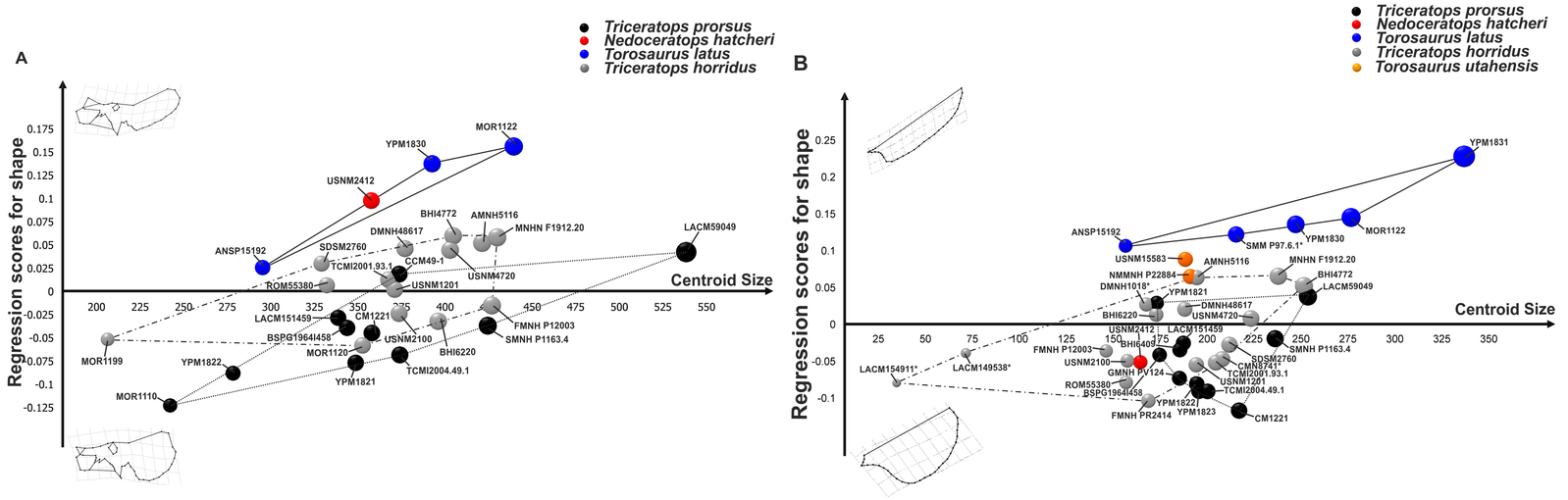
This idea has come under a fair amount of scrutiny, most notably by Andrew Farke and Nicholas Longrich. They have analyzed features of the skulls of each animal and noticed some differences in the skulls. Even a computer analysis of morphometric statistics showed that there is a significant difference between the species. Also, subtle but important differences between the animals have been noted. For example, Torosaurus is much more rare than Triceratops overall, Triceratops specimens that seem old enough to be mature, slight differences in age and location of each species without exact overlap, and no other known ceratopsians which gain holes in frills with age.
Stygimoloch and Dracorex
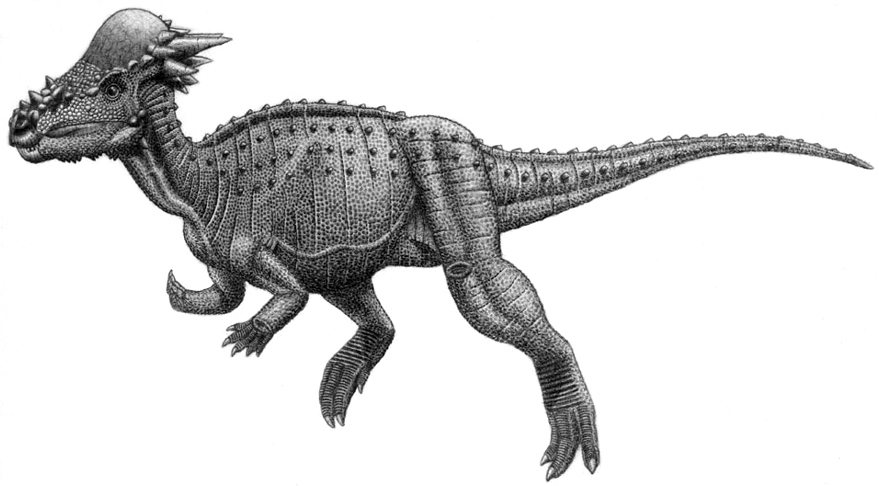
Pachycephalosaurs are dinosaurs famous for their thick skull domes. They are also some of the most poorly understood dinosaurs, as a complete skeleton has never been found of any member of the group. They are also related to the ceratopsians (within Marginocephalia) and have similar growth changes. This had led to significant species controversies.
Pachycephalosaurus, the ‘thick-headed lizard’ as its name describes, was first identified in 1931. In 1983, a relative of Pachycephalosaurus named Stygimoloch (“Demon of the River Styx”) was named, and in 2006, another relative named Dracorex hogwartsia (“Dragon King of Hogwarts”) was named. Just a year later, scientists like Jack Horner had started to question these other species as growth stages of Pachycephalosaurus. In 2009 a study was published by Horner and Goodwin showing that specific points on the skull of each individual species can easily be correlated, supporting the idea that Dracorex being a juvenile, Stygimoloch being a sub adult, and Pachycephalosaurus being an adult and the proper name of the genus. This had spread to other members of the group as well; other flat-headed pachycephalosaurs (like Goyocephale and Homalocephale) have been proposed as juveniles of other genuses.
To Feather or Not To Feather – Dinosaur Body Coverings
Early Feathers
Our interpretations about the coverings of dinosaurs have gone through many iterations over the years. The first dinosaur reconstructions were entirely scaly, in keeping with reptilian ancestry, even with finds like Archaeopteryx pointing to a feathered linage. Early fossil hunters reportedly only cared for the bones, and potentially hundreds of skin and feather impressions were lost to their carelessness. After the dinosaur renaissance and new finds in the later half of the 20th Century, feathers became more accepted. If anything, the feather debate has swung too far, suggesting all dinosaurs were covered in fuzz and fluff. Most dinosaurs have no remains that indicate feathers or not, so plenty is still unknown about dinosaur skin covering. But let us review what is known.

How and why did feathers evolve in the first place? Feathers are not fundamentally different from other vertebrate animal body coverings. The proteins found in feathers, mammal hair, and nails are keratin, similar to what is found in scales of reptiles. This means that feathers can be considered a homologous trait via divergent evolution from a common ancestor with modern reptiles. According to a recent overview of feather evolution (Xu and Guo, 2009), feathers first evolved as a single hair-like object and eventually became more complex. Most scientists infer that feathers would have started out for insulation, and eventually been adapted for service in new functions, such as display and flight. This is corroborated by the fact that smaller dinosaurs have been the most often found with feathers and other coverings. The smaller the animal, the higher the animal’s body surface area to volume ratio, meaning heat loss can occur more quickly. Feathers could reduce this heat loss.

The fossil record does not give us many early examples of feathers, so the initial origin of feathers as a biological structure is hard to know. What is known is that almost all examples of feathers are found on dinosaurs, both avian and non-avian. One possible exception to that rule is found on the enigmatic fossil reptile Longisquama, known from the Triassic of Kazakhstan. Its name, meaning “long scales,” refers to the large feather-like protrusions sticking from the animal’s back. There is much debate over the purpose of these features, or if they are truly feathers in the first place. One animal that Logisquama may be related to is Coelurosauravus, an early reptile which has a membrane protruding from its sides, and presumably used for gliding. Could Logisquama’s long scales be used for gliding, too? Or were they for display to attract mates or identify members of its kind? More specimens and more discoveries of related animals may need to be found before we can know for sure.
Which dinosaurs had feathers?
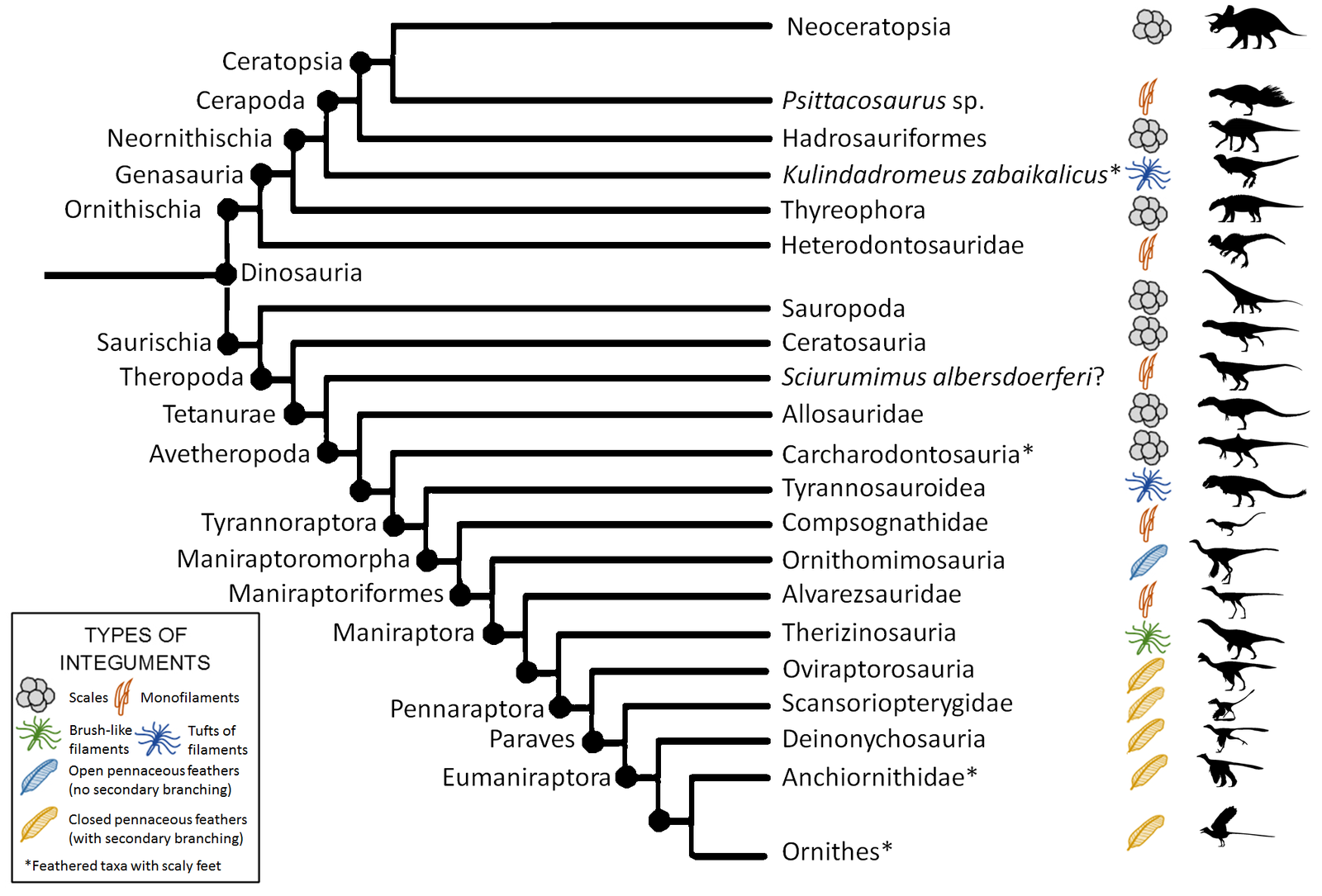
Dinosaurs with feathers or feather-like structures have been found on several branches of the dinosaur family tree. Interestingly, even some ornithischian dinosaurs had feather-like filaments. The pterosaurs had pycnofibres, which were dense bristle-like structures. While these were not exactly feathers, recent studies have shown more variety suggesting that some of these might correspond to dinosaur feathers. Another example, Psittacosaurus, a small early member of the ceratopsian lineage, has been found with a large number of filaments sticking out of its back, from the hip to the mid-tail. Full pennaceous (flying style) feathers, like those seen in modern birds, are found only in maniraptorans, with the most basal example from the ornithomimids. This means that the famous raptors of the Jurassic Park movies would almost certainly have been feathered. In fact, Velociraptor itself has been found with quill knobs, small impressions in the bone of the forearm where a feather would originate. Of course, complex feathers and significant coverings are only found on the coelurosaur branch of the theropods, with some paleontologists thinking all coelurosaurs had feathers.

How about the Jurassic Park co-star, Tyrannosaurus rex? In 2012, a dinosaur named Yutyrannus was described. Meaning “feathered tyrant,” it is the largest dinosaur yet found with evidence of feathers. It has also been phylogenetically placed within the superfamily Tyrannosauridea, and thus could properly be called a tyrannosaur. Tyrannosaurus is considered further down the evolutionary tree with respect to Yutyrannus. Does that mean T. rex had feathers?
The most up-to-date information, based on both T. rex and closely-related large animals, has concluded that the feathers they had were scarce at best. The video game Saurian compiled this information based on published studies to make possibly the most accurate reconstruction yet. Despite this, a 2019 American Museum of Natural History exhibit shows Tyrannosaurus with a feathery body, making the argument that related animals indicate feathers were possible. The exhibit also shows a juvenile with feathers. This is another possibility: even if the adult version was bald, hatchlings may have needed the ‘down’ and could have lost it as they aged.
Dinosaurs to Birds

As evolution progressed, several non-avian dinosaur groups became more and more bird-like, with feathers being only part of the story. This group of animals belongs to the clade Parves, which means “next to birds.” The ornithomimids, for example, derive their name from Ornithomimus, whose name means “bird mimic.” This is due to their toothless beaks and overall body plan that resembled an ostrich. Other dinosaurs, like Microraptor, had an even more bird-like appearance, with long flight-like feathers on its arms… and also its legs! Microraptor was a dromaeosaur, in the same group as other famous animals like Velociraptor and Deinonychus. The dinosaur group that is most closely related to birds but not technically birds are the troodontids. The first non-pterosaur that could have flown was Archaeopteryx. It is part of a group called Avialae, “bird wings.” This is the group where the line between non-avian dinosaurs and birds becomes blurry. Euornithies is the clade that contains true birds, and a subgroup of that is called Aves, the crown group of modern birds.

One of the biggest ongoing debates in bird evolution is how flight evolved itself. There are two main camps on this question. One idea is that feathers enabled these theropods to be more agile on the ground. As their agility increased, feather complexity also increased, eventually aiding their flight. The other idea proposes these animals already lived in trees, and were gliding from branch to branch. Their feathers assisted in gliding, and eventually led to flight. There is also an in-between, in which the wings helped with climbing, as some modern birds have been experimentally shown to do today. There are compelling arguments on all sides, so no one explanation has yet achieved widespread acceptance.
Movement and thermoregulation – Get that blood flowing!

How dinosaurs moved and how dinosaurs regulated their internal temperature has been debated and controversial since their discovery. What may be less obvious is how much these ideas are interlinked. Dinosaurs’ closest living relatives, crocodiles, are ectothermic (commonly known as cold blooded). This means they rely on their environment for warmth. This has advantages and disadvantages: less resources are needed, but they are less adaptable to cold climates and have limitations on endurance. Crocodiles, by the way, have several advantages over their ectothermic kin. They have a 4-chambered heart for better blood exchange efficiency, a secondary palate to allow breathing while eating, and the ability to walk upright (known as a high walk) for more efficiency in movement. These are adaptations that mammals share, and most scientists think dinosaurs had these adaptations as well.

Birds, the direct descendants of dinosaurs, are mostly endothermic (commonly known as warm blooded). That means birds (and the other endotherms, mammals) can live in many more climates across Earth and have a higher metabolism. This allows for faster movement but also requires more resources. It should be noted that there are also mesotherms, animals that are not quite warm blooded or cold blooded, but in between. Another complexity is homeothermy, the ability to maintain body temperature so that metabolic processes can be maintained. This is related to endothermy, but is distinct, because some endotherms do not have homeothermy. The question remains: where do dinosaurs fit into this spectrum? And how do we find out?
Early dinosaur movement

When dinosaurs were first described, they were thought to be related to modern reptiles, and were imagined as just larger versions of them. They were slow and lumbering in the minds of scientists at the time and were assumed to be ectothermic as well. They were mounted with sprawled legs and dragging tails. Evidence started to build that dinosaurs were related but distinct from living reptiles on Earth today. For example, dinosaurs have their legs positioned under their bodies. This is not just an aesthetic choice; a sprawled posture is much more energy intensive, so dinosaurs were more efficient than their reptile brethren. Also, sprawled posture interferes with breathing by flexing the lungs (called Carrier’s Constraint), causing most sprawled animals to hold their breath while moving. Pontzer et al. (2009) argued that the posture of dinosaurs links them to endothermy alone. Even in 1869, Thomas Henry Huxley had concluded that dinosaurs were more closely related to birds as part of his defense of the newly-proposed theory of evolution by Charles Darwin. These ideas did not help the image of dinosaurs in the minds of the public, however. Even into the mid 20th Century, dinosaurs were nothing but big cold-blooded lizards.
Dinosaur Renaissance

Starting in 1964, John Ostrom, his student Robert Bakker, and others discovered and described the animal Deinonychus. With its small, light frame and bird-like posture, it was a great example that went against the slow and sluggish stereotype dinosaurs had in public opinion. This started what is now known as the Dinosaur Renaissance. Ideas that changed during this time include: a closer evolutionary connection to birds, a more nimble posture (without dragging tails), stronger ideas about herd behavior, and a more open mind on endothermic thermoregulation. The current idea about the extinction of the non-avian dinosaurs by a meteor impact also was developed during this time.

This revolution in the thinking about dinosaurs has basically continued to this day, but the debate over dinosaur thermoregulation is still ongoing, mostly because it is harder to decipher than some of the other debates. One practice that has increased our understanding of physiology of dinosaurs is histology: the microscopic study of tissues, namely bone. Several studies have found that dinosaurs had fast growth rates. They also had fibrolamellar bone and Haversian canals, both of which are common in endothermic animals (but not exclusive to them).
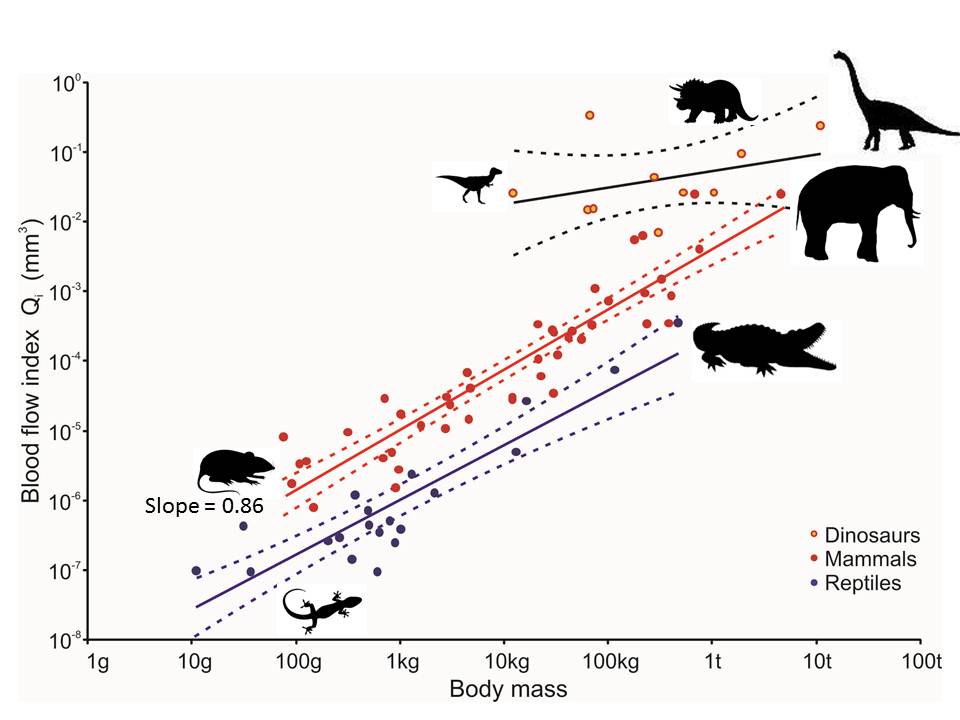
Bone structures can also estimate blood flow, with high blood flow being attributed to endothermy. By looking at the small holes throughout bones, called nutrient foramina, and comparing the size of the holes to the size of the animal, you can estimate whether the animal is endothermic or ectothermic. As you can see from the figure, dinosaurs match much more closely with mammals, and may have been highly active, and possible endotherms as a result.
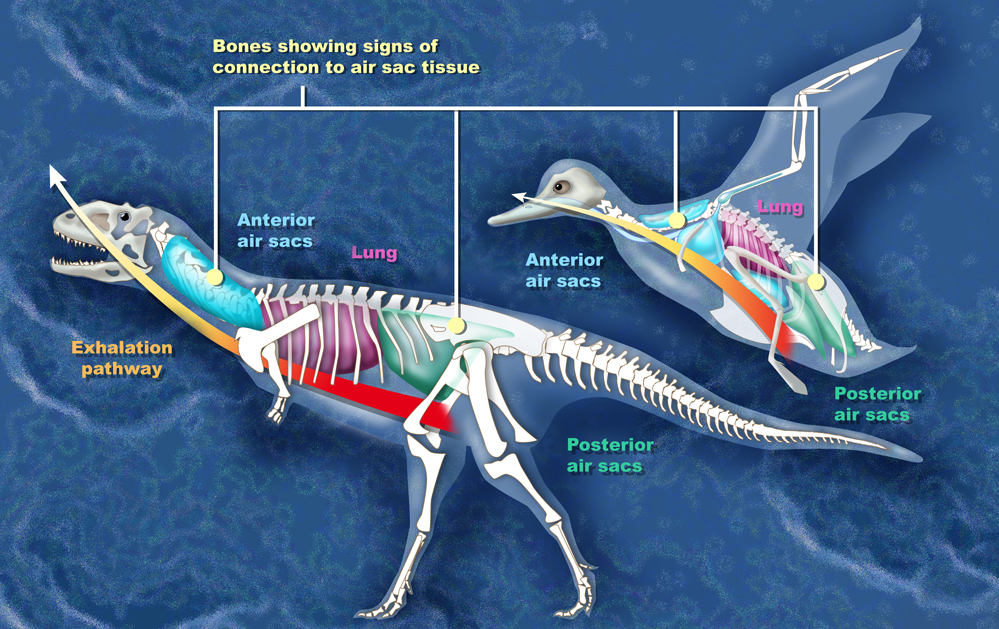
Another clue comes from breathing. Some scientists think dinosaurs had an air sac system similar to birds. The sacs are found mostly in their skulls and vertebrae, and would have allowed more efficient exchange of oxygen. They have also been pointed to as a weight-saving system in larger species. Of course, these features are almost impossible to fossilize. While there is no doubt that dinosaur descendants (birds) have them, where they developed in dinosaur’s evolutionary history is very much up for debate. Theropods, the branch of the dinosaur tree that turned into birds, have the best evidence for possible air sacs in non-avian dinosaurs.
Endotherm without Endothermy?

The paragraphs above were just a partial list of evidence that could be used to explain if dinosaurs were endothermic or ectothermic, there is another factor to consider: dinosaurs could have been functionally endotherms without being actual endotherms. How? By keeping in their heat. This could have happened in two main ways. First, feathers (or similar proto-feather structures) may have evolved as a warmth structure first, then became used for flight later. Secondly is an idea that is called inertial endothermy, bulk endothermy, or gigantothermy. This suggests the large size of dinosaurs, with selective blood pumping taking advantage of surface area to volume ratio, could have had high blood temperatures without the food costs of endothermy. Of course, smaller dinosaurs and the young of larger dinosaurs would not have had this advantage, but feathers on the smaller animals or young could have helped.
Dino Debate – Did I Get It?
Further Reading
“New Analyses Of Dinosaur Growth May Wipe Out One-third Of Species”. Sciencedaily.com. 2009-10-31. Retrieved 2010-08-03.
Day, Julia J.; Upchurch, Paul; Norman, David B.; et al. (May 31, 2002). “Sauropod Trackways, Evolution, and Behavior”. Science. Washington, D.C.: American Association for the Advancement of Science. 296 (5573): 1659. doi:10.1126/science.1070167. ISSN 0036-8075. PMID 12040187.
Horner J.R. and Goodwin, M.B. (2009). “Extreme cranial ontogeny in the Upper Cretaceous Dinosaur Pachycephalosaurus.” PLoS ONE, 4(10): e7626. (Online full text)
Horner, J.R.; Makela, R. (1979). “Nest of juveniles provides evidence of family structure among dinosaurs”. Nature. 282 (5736): 296–298. Bibcode:1979Natur.282..296H. doi:10.1038/282296a0.
Longrich, N. R. & Field, D. J. (2012). “Torosaurus is not Triceratops: Ontogeny in chasmosaurine ceratopsids as a case study in dinosaur taxonomy”. PLoS ONE. 7 (2): e32623. Bibcode:2012PLoSO…732623L. doi:10.1371/journal.pone.0032623. PMC 3290593. PMID 22393425.
Maiorino L., Farke A.A., Kotsakis T., Piras P.,2013, “Is TorosaurusTriceratops? Geometric Morphometric Evidence of Late Maastrichtian Ceratopsid Dinosaurs”, PLoS ONE 8(11): e81608. doi:10.1371/journal.pone.0081608
Pontzer, H. .; Allen, V. .; Hutchinson, J. .; Farke, A. A. (2009). Farke, Andrew Allen (ed.). “Biomechanics of running indicates endothermy in bipedal dinosaurs”. PLOS ONE. 4 (11): e7783. Bibcode:2009PLoSO…4.7783P. doi:10.1371/journal.pone.0007783. PMC 2772121. PMID 19911059.
Scannella, J. and Horner, J.R. (2010). “Torosaurus Marsh, 1891, is Triceratops Marsh, 1889 (Ceratopsidae: Chasmosaurinae): synonymy through ontogeny .” Journal of Vertebrate Paleontology, 30(4): 1157–1168.
Seymour, Roger S.; Smith, Sarah L; White, Craig R.; Henderson, Donald M.; Schwarz-Wings, Daniela (2012). “Blood flow to long bones indicates activity metabolism in mammals, reptiles and dinosaurs”. Proceedings of the Royal Society B. 279 (1728): 451–456. doi:10.1098/rspb.2011.0968. PMC 3234558. PMID 21733896
Switek, Brian. “New Study Says Torosaurus=Triceratops”. Dinosaur Tracking. Smithsonian.com. Retrieved 2 March 2011.